On the Origins of New Forms of Life
9.11: Proboscideans (Elephants)
EUGENE M. MCCARTHY, PHD GENETICS
|
|
(Continued from the previous page)

|
|
Florentino Ameghino (1865-1936) |

|
|
Carlos Ameghino (1854-1911) |
During the late 1800s Florentino and Carlos Ameghino discovered in South America the remains of a huge extinct mammal. They named it Pyrotherium or "fire beast," a reference to the volcanic ash in which the first specimens were found. Pyrotherium and related forms ("pyrotheriids") were a long-lived group prominent in South American deposits from the late Paleocene to Upper Miocene.1 The Ameghinos believed that Pyrotherium was an early proboscidean.2
Amherst professor Frederic B. Loomis (1914) discovered an intact skull of Pyrotherium, which seemed to him clearly to demonstrate its proboscidean affinity (see Loomis, p. 162, where he places Pyrotherium in Proboscidea).
Romer (1966: 246) points out that Pyrotherium was not only
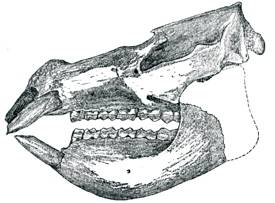 Skull of Pyrotherium
Length: 38 in (96.5 cm)
From Loomis (1913: 136)
Skull of Pyrotherium
Length: 38 in (96.5 cm)
From Loomis (1913: 136)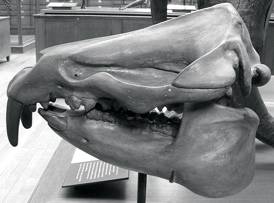 Moeritherium, an acknowledged early proboscidean.
Muséum national d'Histoire naturelle, Paris
Moeritherium, an acknowledged early proboscidean.
Muséum national d'Histoire naturelle, Paris
Artist: Heinrich Harder
But for many biologists similarity — or, seemingly, even identity — is not enough. In the same place, Romer goes on to express his belief that these resemblances between pyrotheres and the early proboscideans of Africa "are surely a case of exceedingly close parallelism." In Romer's defense, it can be said that he was writing at a time when continental drift was not yet an established geophysical phenomenon. He thought in terms of fixed continents and believed elephants (and all other placental mammals) had rapidly evolved from small, generalized late Cretaceous insectivores. As such, his perspective excluded the idea that pyrotheres might be connected with elephants in other parts of the globe (but even some early writers thought the similarity of Pyrotherium and Barytherium indicated a "land bridge" had once existed between Africa and South America3). Such similarities extend to the various features of the skeleton. For example, Salas et al. (2006: 765) point out
Dixon et al. (1966: 249) also note the extreme similarity between pyrotheres and elephants:
But these authors, like Romer, attribute the similarity not to genetic relationship, but to convergent evolution.
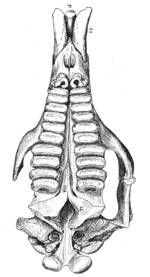 Pyrotherium - upper teeth (Loomis 1914)
Pyrotherium - upper teeth (Loomis 1914)
Barytherium is known from the late Eocene and early Oligocene of North Africa.4 Dixon and his colleagues appear to be correct in asserting that Barytherium and Pyrotherium were similar. Comparison reveals their teeth are virtually identical.5 These two animals both had bilophodont teeth (both premolars and molars) of the same size and shape. In both, the second upper, and first lower incisors formed chisel-shaped tusks bearing enamel only on the outer surface of the tooth. The claim that "convergent evolution" could produce such extreme similarity is implausible.
Evidence discussed thus far (in Section 9) indicates a much earlier origin for placental mammals than is commonly received. This conclusion, together with our now-firm knowledge that Africa and South America were in direct contact in the early Cretaceous, suggests pyrotheriids and barytheriids are not just forms that evolved in "parallel." It is more plausible to suppose that Pyrotherium, and related South American forms, were forms descended the same ancestral proboscidean similarity set giving rise to Barytherium in Africa (and that that set existed prior to the separation of those two continents). In other words, it seems likely proboscideans were already in existence in Gondwana in the early Cretaceous. Thus, proboscideans would be many millions of years older than biologists generally believe. In fact, it is even conceivable that further analysis of fossils will show Pyrotherium and Barytherium are merely distinct names applied to a single form that predated and long survived the separation of Africa and South America.6
Notes (Works Cited):
1.Benton (1993); Romer (1966); Savage and Russell (1983).
2.Ameghino (1889, 1895, 1902). Lydekker (1896: 85) also identified Pyrotherium as an elephant. See also: Andrews (1904: 101).
3.For example, Andrews (1904: 101).
4.Savage and Russell (1983: 114); Shoshani and Tassy (1996: 64).
5.MacFadden and Frailey (1984, Fig. 2 & Table 1); Shoshani and Tassy (1996, Plate 8.3).
6.Another form worthy of investigation with regard to proboscideans is Diprotodon of the Australian Pleistocene. This large animal has sometimes been described as a marsupial elephant. However, its soft anatomy is unknown, and its skeleton bears little if any resemblance to those of extant marsupials (the discussion of marsupials explains why extinct forms cannot be diagnosed as marsupials).
9.11: Proboscideans (Elephants)- © Macroevolution.net
8.1: Dubious Assumptions - © Macroevolution.net