The Hybrid Hypothesis
1: Human origins: Are we hybrids?
|
Die Stimme der Vernunft ist leise.
—Sigmund Freud
|
Ballarat, Victoria, Australia
|
In a Los Angeles Review of Books article longtime science journalist Greg Critser examines the hybrid theory of human origins.
|
This section is a little different from others on this site, because it’s about the findings of my own research. I am a University of Georgia trained geneticist (M.S., Ph.D.) who worked in various genetics laboratories at the University of Georgia and conducted research there from 1989 to 2007. During those years I also taught biology and genetics at UGA.
My work focuses on hybrids and, particularly, the role of hybridization in the evolutionary process. Here, I report certain facts, which seem to indicate that human origins can be traced to hybridization, specifically to hybridization involving the chimpanzee (but not the kind of hybridization you might suppose!). You can access detailed and documented discussions supporting this claim from the table of contents. But I’ll summarize the basic reasoning here, without a lot of citations and footnotes (see below).
My theory of human origins: Rationale
|
Always remember that it is impossible to speak in such a way that you cannot be misunderstood.
—Karl Popper
|
|
From an article on mules elsewhere on this website: “Zirkle (1935, p. 7) says that the sterility of the mule made it the first animal whose hybrid origin was generally recognized because “the origin of fertile hybrids could easily be forgotten, particularly the origin of those which appeared before the dawn of history.” The mule, however, was so sterile that it was necessary to produce it with the original cross (Zirkle provides extensive information on the early history of mule). The fact that the hybrid origin of the mule has so long been known, together with its marked sterility, has no doubt greatly contributed to the widespread, but erroneous belief that all hybrids are sterile.”
More about mules >> |
So why do I think humans are hybrids? Well, first of all, I’ve had a different experience from most people. I’ve spent most of my life (the last thirty years) studying hybrids, particularly avian and mammalian hybrids. I’ve read thousands of reports describing them. And this experience has dispelled some mistaken ideas I once had about hybrids, notions that I think many other people continue to take for granted.
For example, one widespread, but erroneous, belief is that all hybrids are sterile. This idea keeps a lot of people from even considering the possibility that humans might be of hybrid origin. The reality, however, is something quite different. For instance, in reviewing the reports I collected for my book on hybridization in birds (Handbook of Avian Hybrids of the World, Oxford University Press, 2006), which documents some 4,000 different kinds of hybrid crosses among birds, I found that those crosses producing partially fertile hybrids are about eight times as common as crosses known to produce sterile ones. The usual result is a reduction in fertility, not absolute sterility. My current work documenting hybridization among mammals shows that partially fertile natural hybrids are common, too, in Class Mammalia. And yet, it seems most people base their ideas of hybrids on the common mule (horse x ass), which is an exceptionally sterile hybrid, and not at all representative of hybrids as a whole.
I should, perhaps, also mention that differences in parental chromosome counts, even rather large ones, do not preclude the production of fertile hybrids. While differences of this sort do bode ill for the fertility of the resulting progeny, it is only a rule of thumb. For example, female geeps, the products of hybridization between sheep (2n=54) and goats (2n=60), can produce offspring in backcrosses. Likewise, female zeedonks (Burchell’s Zebra, 2n=44 x Ass, 2n=62) have also been fertile in backcrosses. There are many other examples of this sort among mammalian hybrids. Therefore, such differences between the parents in a cross do not in any way guarantee an absolute sterility in the hybrid offspring. (For those readers who do not know, backcross hybrids are produced when hybrids from a first cross mate with either of the two types of parents that produced them. When the resulting progeny mate again with the same parental type, the result is the second backcross generation, and so forth.)
Dear Gene,
Thank you again, I've read your profound considerations with the greatest interest. I think nowadays the situation is worse than it used to be at Darwin’s age. The case of physics (cosmology and high energy physics, above all) is emblematic: big science, massive investments involving thousands of researchers, industrial operations essentially leading nowhere. New ideas cannot filter through, they would too badly damage ongoing business. Even people iconized in the first part of their life like Fred Hoyle, Dirac, Einstein have been in the latter part considered kind of nuts since they didn't abjure independent thinking to adore any ‘standard model’ idol, like present-day believers.
Your proposal in this environment is very courageous, and the huge and admirable work you carried out to supplement and document it vigorously goes in the direction of re-establishing sensible science in one of the most relevant subjects for human society.
The aim surely deserves the effort.
|
Dear Dr. McCarthy, I admire your work as you described in your recent newsletter. I was surprised by the reaction of the chattering biologic community and was pleased by your calm reaction to it all. Now I know what Darwin went through. You have my support and if this old biologist/physician/urologist can help, please let me know. I've directed all my bio friends to your site.
—An email from a supporter
|
A second so-called fact, which might make it seem impossible for humans to have had a hybrid origin, is the equally erroneous notion that hybrids, especially successful hybrids, do not occur in a state of nature. A third is the mistaken idea that only plants hybridize, and never animals. In fact, however, natural, viable, fertile animal hybrids are abundant. A wide variety of such hybrids occur on an ongoing basis (read a detailed discussion documenting these facts). For example, of the more than 4,000 different types of hybrid crosses listed in my book on hybridization in birds, approximately half are known to occur in a natural setting (download a PowerPoint presentation summarizing data on hybridization in birds). My current research indicates a comparable rate for mammals.
Sequence data. And I must now emphasize a fact that I, as a geneticist, find somewhat disappointing: Though there are other ways of detecting them, with nucleotide sequence data, it can be very difficult to identify later-generation backcross hybrids derived from several repeated generations of backcrossing (Vähä and Primmer 2006, Engler et al. 2015). And this would be especially true of any remote descendants of backcross hybrids produced in ancient times, which is what I'm proposing humans may actually be. To better understand why backcross hybrids are hard to analyze with sequence data, take a look at this diagram:
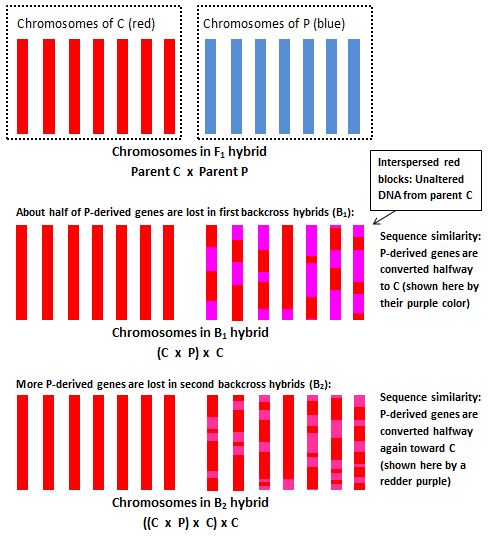
The following rebuttals address the most common objections raised against the hybrid theory of human origins:
Instead, as is the case with other later-generation backcross hybrids, the most revealing data is of an anatomical and/or physiological nature. And this is exactly the kind of hybrid that humans seem to be, that is, it appears that humans are the result of multiple generations of backcrossing to the chimpanzee.
The thing that makes backcross hybrids hard to analyze using genetic techniques is that, in terms of nucleotide sequences, they can differ very little from the parent to which backcrossing occurs. It’s important to realize, however, that a lack of such differences does not prevent them from differing anatomically. Sequence differences are not necessary for anatomical differences to be present. An obvious example of this phenomenon is Down’s syndrome. Individuals affected by Down’s regularly exhibit certain distinctive anatomical features, and yet in terms of their nucleotide sequences they do not differ in any way from other humans. To detect someone with Down’s syndrome, sequence data is completely useless. But with anatomical data, detecting affected individuals is easy. This issue is discussed in more detail in a subsequent section. The key fact is that with Down’s syndrome the differences that we see are due to differences in the number of genes present, that is, dosage differences, and not to differences in the nucleotide sequences of those genes. Dosage differences of this sort are exactly what hybridization typically produces.
Human infertility. Another observation that appears significant in connection with the theory of human origins here under consideration is that it has been well known for decades that human sperm is abnormal in comparison with that of the typical mammal. Human spermatozoa are not of one uniform type as in the vast majority of all other types of animals. Moreover, human sperm is not merely abnormal in appearance — a high percentage of human spermatozoa are actually dysfunctional. These and other facts demonstrate that human fertility is low in comparison with that of other mammals (for detailed documentation of this fact see the article Evidence of Human Infertility). Infertility and sperm abnormalities are characteristic of hybrids. So this finding suggests that it's reasonable to suppose, at least for the sake of argument, that human origins can be traced to a hybrid cross. It is also consistent with the idea that the hybridization in question was between two rather distinct and genetically incompatible types of animals, that is, it was a distant cross.
Methodology. The chimpanzee is plausible in the role of one of the parents that crossed to produce the human race because they are generally recognized as being closest to humans in terms of their genetics (here, I use the term chimpanzee loosely to refer to either the common chimpanzee or to the bonobo, also known as the pygmy chimpanzee; the specific roles of these two rather similar apes within the context of the theory of human origins now under consideration will be explained in a subsequent section). But then the question arises: If an ancient cross between the chimpanzee and some parental form “X” produced the first humans, then what was that parent? Does it still exist? What was it like?
As the reader might imagine, if the assumption is correct that one of our parents is the chimpanzee, then it should be possible actually to identify the other parent as well. A hybrid combines traits otherwise seen only separately in the two parental forms from which it is derived, and it is typically intermediate to those parents with respect to a wide range of characters. Naturalists routinely use these facts to identify the parents of hybrids of unknown origin, even backcross hybrids.
First they posit a particular type of organism as similar to the putative hybrid (in the present case, this organism is the chimpanzee). They then list traits distinguishing the hybrid from the hypothesized parent, and this list of distinguishing traits will describe the second parent. A detailed analysis of such a triad will often establish the parentage of the hybrid. The traits in question in such studies are generally anatomical, not genetic. DNA evidence is used in only a very small percentage of such identifications (and even then, rarely in efforts to identify backcross hybrids), and yet firm conclusions can generally be reached.
So in the specific case of humans, if the two assumptions made thus far are correct (i.e., (1) that humans actually are hybrids, and (2) that the chimpanzee actually is one of our two parents), then a list of traits distinguishing human beings from chimpanzees should describe the other parent involved in the cross. And by applying this sort of methodology, I did in fact succeed in narrowing things down to a particular candidate. That is, I looked up every human distinction that I could find and, so long as it was cited by an expert (physical anthropologist, anatomist, etc.), I put it on a list. And that list (see next page), which includes many traits, consistently describes a particular animal. Keep reading and I’ll explain. Next page >>
|
The scientist has a lot of experience with ignorance and doubt and uncertainty, and this experience is of very great importance, I think. When a scientist doesn’t know the answer to a problem, he is ignorant. When he has a hunch as to what the result is, he is uncertain. And when he is pretty darn sure of what the result is going to be, he is in some doubt. We have found it of paramount importance that in order to progress we must recognize the ignorance and leave room for doubt. Scientific knowledge is a body of statements of varying degrees of certainty — some most unsure, some nearly sure, none absolutely certain.
—Richard Feynman
|