On the Origins of New Forms of Life
8.2: Similarity Chains and Similarity Sets
EUGENE M. MCCARTHY, PHD GENETICS
|
|
(Continued from the previous page)
Intermediate forms can be used to unite disparate types into similarity chains in which those types that form adjacent links in the chain are relatively similar. Although those types at the two ends of the chain might be quite different, forms constituting adjacent links in such a chain might plausibly produce new types by stabilization processes involving hybridization. For example, most people would think that a mallard duck (Anas platyrhynchos) would be unable to hybridize with a mute swan (Cygnus olor). Indeed, no such hybrids seem to have been reported. However, both of these birds do hybridize with the Canada Goose (Branta canadensis) and hybrids of both types have been reported from a natural setting (pictures). Among birds as a whole, the author has found that such chains of hybridizing forms can be much longer. For example, Figure 8.1, which summarizes reports of hybridization among upland game birds, clearly shows that a wide variety of similarity chains could be constructed among such birds.
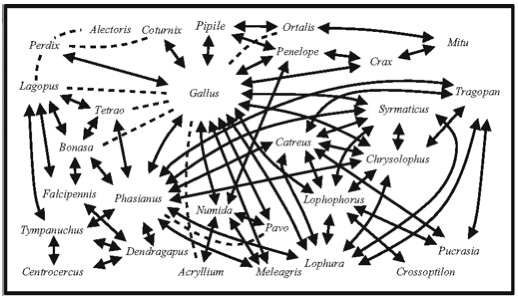 Figure 8.1. Diagrammatic representation of reported hybridization among upland game bird genera (arrows connecting genus names indicate hybridization has been reported between the two connected genera; dotted lines indicate that available reports need further verification). From McCarthy (2006).
Figure 8.1. Diagrammatic representation of reported hybridization among upland game bird genera (arrows connecting genus names indicate hybridization has been reported between the two connected genera; dotted lines indicate that available reports need further verification). From McCarthy (2006).
This idea of chains of hybridizing forms is a fundamental principle in stabilization theory. Over evolutionary time, the composition of such chains would change as (1) new forms of life occasionally came into being; and (2) older forms occasionally died out. A trait initially present in only a single form in an ancient chain might therefore spread with time until it was present in many or all forms in a modern chain. The trait might spread merely at random, but it would spread more rapidly if those forms having it could better avoid extinction. For example, early plants lacked vascular systems. But suppose a form in an early similarity chain (ancestral to modern vascular plants) obtained this trait (or an approximation of it). Such a trait would be advantageous and would permit its possessor to exploit drier environments inaccessible to other types of organisms in the same similarity chain. In successive chains more and more new vascular forms would appear because vascular forms would be fitter than nonvascular forms. They would thus exist for a longer time and produce more offspring forms than nonvascular forms. When new forms are produced via stabilization processes involving hybridization between adjacent forms in the chain, vascular forms would come to predominate, as is the case with modern plants. Thinking in terms of similarity chains, then, one can see how an increasing number of forms with a particular trait can come into being even when the trait itself originally came into being only once in a single type of organism. The general nature of this process is diagrammed in Figure 8.2.
One can also see why the potential power of point mutation is greatly amplified under stabilization theory. For by spreading in successive chains from the form that initially obtained it, such a mutation can affect a much broader range of organisms. Moreover, each individual organism has far more ancestors in which such a point mutation can occur (see Figure 7.1). Under this view, similarity chains succeed each other in a manner parallel to the succession of generations in an ordinary population. Note that the parent forms and offspring forms in this process are analogous to the parent individuals and offspring individuals of neo-Darwinian theory. Those parent forms with traits favored by natural selection produce more offspring forms (whereas under neo-Darwinian theory, those parent individuals with advantageous traits produce more offspring individuals). The selection is between types of organisms, not individuals. There is no slow change of one form into another. There is only production of new stable forms by preexisting stable forms.
Similarity chains are really an oversimplification. Normally the ability to hybridize is not limited to a set of organisms that can be arranged in a simple linear chain. Many forms treated as species have the ability to hybridize with numerous partner forms. For example, the mallard duck (Anas platyrhynchos) has been reported to hybridize with about 60 other waterfowl treated as species. More realistically, then, one can posit a similarity set, a set of forms in which all members are connected to at least one other member by the potential to hybridize. In such a set, a favorable trait would be able to spread to an increasing number of set members over time. The situation would then be analogous to neo-Darwinism's description of the spread of a favorable trait within an ordinary population (the set of individuals making up a population is analogous to the set of forms making up a similarity set).
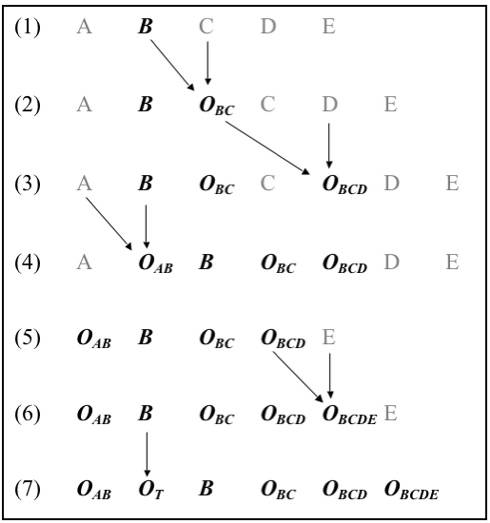 Figure 8.2. A series of similarity chains showing the spread over time of an advantageous trait (forms with this trait are shown in bold italic); (1) Initial similarity chain. A series of five different types of organisms, A, B, C, D, E (adjacent forms in the series are assumed to be more similar and hence more hybridizable). Initially, the only form with the advantageous trait is B; (2) B has hybridized with C to produce offspring form OBC (in the figure "O" stands for "offspring form"); (3) OBC has hybridized with D to produce offspring form OBCD; (4) A has hybridized with B to produce offspring form OAB; C has become extinct; (5) A and D have become extinct; (6) OBCD has hybridized with E to produce offspring form OBCDE; (7) B has produced a tetraploid OT; E has gone extinct. From the set of forms in the original chain only B has survived; all forms in the similarity chain now have the trait.
Figure 8.2. A series of similarity chains showing the spread over time of an advantageous trait (forms with this trait are shown in bold italic); (1) Initial similarity chain. A series of five different types of organisms, A, B, C, D, E (adjacent forms in the series are assumed to be more similar and hence more hybridizable). Initially, the only form with the advantageous trait is B; (2) B has hybridized with C to produce offspring form OBC (in the figure "O" stands for "offspring form"); (3) OBC has hybridized with D to produce offspring form OBCD; (4) A has hybridized with B to produce offspring form OAB; C has become extinct; (5) A and D have become extinct; (6) OBCD has hybridized with E to produce offspring form OBCDE; (7) B has produced a tetraploid OT; E has gone extinct. From the set of forms in the original chain only B has survived; all forms in the similarity chain now have the trait.
The reader might correctly suppose the large differences between some types of organisms prevent the production of hybrids. This is indeed the case. However, such gaps may have been bridged in former times by extinct (stable and discrete) intermediate forms, so that hybridizing forms would be less disparate. It is also possible hybridization could in some cases occur despite our perception of a large disparity between the taxa involved. Indeed, broad surveys of hybridization in birds and mammals (McCarthy 2006; McCarthy, this website) indicate similarity, whether genetic or morphological, is an imperfect predictor of crossability.
Moreover, intermediate forms often connect taxonomic categories supposed to be entirely distinct. In the previous section, it was pointed out that intermediate forms connect even the broadest categories (plants, animals, fungi, bacteria). It is not surprising, then, that lesser categories are connected as well. For example, among mammals, flying lemurs (genus Cynocephalus) are often placed in a separate order of their own (Dermoptera), but various authors have also classified them on various occasions as bats (Order Chiroptera), primates (Order Primates), and insectivores (Order Insectivora). For example, Duff and Lawson (2004), the taxonomic standard used for mammals discussed in this book, classify them as primates. The Raccoon Dog (Nyctereutes procyonoides) is placed in the same family as dogs (Canidae), but it is obviously similar to raccoons, which belong to a different family (Procyonidae). Scientists have long argued over whether tree shrews (Tupaiidae) are primates or insectivores. Classified as a cat (Family Felidae), the Jaguarundi (Felis yagouaroundi) resembles a weasel or otter (Family Mustelidae), while the Fossa (Cryptoprocta ferox) seems to connect the cat family with the civets and genets (Family Viverridae). Classified as a carnivore (Order Carnivora), the kinkajou (Potos flavus) is very similar to a primate. The clear-winged moths, classified as Family Sesiidae of Order Lepidoptera, have a variety of features that are hornet- or beelike. Both bees and hornets are classified as belonging to a separate order (Hymenoptera). Bivalved gastropods (Family Juliidae) are intermediate between snails and bivalved molluscs, normally treated as separate classes (Gastropoda and Bivalvia) of the phylum Mollusca. In other words, juliids bridge the gap between categories that, from a taxonomic standpoint, are as distinct as a bird and a fish. Considered even more distinct (different phyla), annelid worms and arthropods are connected by onychophorans, which have traits characteristic of both. One could go on and on. Such forms seem innumerable. Once again, Table 8.1 gives a (by no means complete) sampling of such organisms. The existence of such intermediates suggests similarity chains may stretch, or formerly stretched, further than one might at first suppose.
Under stabilization theory intermediate organisms are expected. But under neo-Darwinian theory they constitute a problem. For only some of these intermediates can be explained. One of the most common explanations offered under that view is that the form in question belongs to one category but gains an advantage by mimicking another. For example, one might suppose clear-winged moths gain protection from predators by imitating bees and hornets, which also have clear wings. But in many cases explanations in terms of mimicry are subject to obvious objections. Bivalved gastropods have a small, coiled structure like a snail shell attached to the apex of a bivalved shell. They also have heads like those of snails (not found in ordinary bivalves). What is the advantage of a snail mimicking a bivalve, or a bivalve mimicking a snail?
Admittedly, some taxonomic categories seem entirely discrete. They do not seem to be connected by any intermediate forms of any kind. But stabilization theory interprets these well-defined categories in a different way. Under this view, two discretely different but related extant categories are seen as two similarity sets descended from a single ancient similarity set that did contain intermediate forms now extinct. And, in fact, extinct fossil forms do connect many similarity sets that seem to be unconnected today (see Table 8.1). A well-known example is the reptilelike fossil bird Archaeopteryx.
Note that this process of breaking similarity sets into isolated subsets is analogous to the process of divergence described in neo-Darwinian theory. But this sort of divergence involves the extinction of intermediate forms (Figure 8.3), whereas neo-Darwinian divergence involves a dying out of intermediate individuals. Note also that the natural selection among the various forms within an isolated similarity set is analogous to the selection the neo-Darwinian paradigm describes as occurring in a reproductively isolated lineage composed of interbreeding individuals. Note also that the different forms within a similarity set need not correspond to types treated as species. They could just as well be forms treated as genera or families (and in fact, many intergeneric and interfamilial hybrids are known).
Within this context, one can clearly see the significance of the contrasting phenomena of hybrid vigor and hybrid inviability (neo-Darwinists often discuss the latter of these two phenomena under the heading of "hybrid breakdown"). As we have already seen (sections 2, 3, and 4), breeders often observe that the products of particular crosses are more vigorous than, and have characteristics superior to those of, their parents. Nevertheless, evolutionary biologists often discount the creative powers of hybridization by noting that hybrids are frequently inviable, and that inviable hybrids are unlikely to produce a new, successful form. However, crosses of both types do exist, those that produce viable hybrids and those that produce inviable ones. From the perspective of stabilization theory, the implication of this fact is that some crosses will give rise to forms that can outcompete forms produced by other crosses. The propagation of new forms via hybridization, then, is blocked only in the case of those crosses producing inviable hybrids, not those that produce vigorous hybrids with advantageous characteristics. In other words, there is natural selection for certain hybrid forms and against others.
It is well known that many types of organisms today represented by one or a few forms were anciently represented by a diverse array of forms. For example, shelled cephalopods were abundant in the time of the dinosaurs and occurred in a diverse array of forms. But today the Chambered Nautilus ( Nautilus pompilius) and a few closely related forms are the sole survivors. The surviving representatives of such groups are often much smaller than some members of the ancient group. A nautilus is about the size of a dinner plate, but some ancient shelled cephalopods were the size of airplane tires. Modern lycopods (clubmosses) are tiny, but in the Palaeozoic many lycopods were the size of trees. This residual dwarfism is apparent when various modern taxa are compared to their forebears. It may often be an artifact of preservation or simply a result of impression. For example, if forms in a particular category varied widely in size, the largest forms might be preserved as fossils with higher probability or make a greater impression on the observer than the smallest. Under such circumstances a surviving intermediate-sized form might seem like a dwarf, even though smaller forms once existed. Under stabilization theory this sort of reduction in diversity, whether the single remaining form is dwarf or not, is an evolutionary dead-end. The residual form has become isolated. There are no similar types with which it can interact genetically via hybridization. This situation results in an inability, or at least a marked reduction in the capacity, of the isolated form to produce new forms. This type of reproductive isolation would prevent any further evolution from occurring (neo-Darwinian theory describes an analogous situation in which a population can lose much of its ability to evolve due a loss of genetic diversity).
Note that under the view of evolution just described the standard system of taxonomic classification becomes an artificial construct. Its dichotomously branching topology would no longer correspond to a topology of historical events. From the standpoint of stabilization theory, there is really no reason even to have a formal hierarchy of categories. In fact, the niggling assiduity lavished on the classification of each and every type of organism under the present system becomes absurd (e.g., "Kingdom: Animalia; Phylum: Chordata; Class: Mammalia; Order Primates; Family: Hominidae; Subfamily: Homininae; Tribe: Hominini; Genus: Homo; Species: sapiens"). Surely we will be able to find a less cumbersome way of designating a particular sort of organism. For my own part, I would be satisfied to refer to organisms by their common names and to have an identification number that could be added in those cases where any doubts might arise concerning the exact identity of the organism in question. For example, one might write "Bengal Tiger (AGK-345-821)." This format would be compact, unambiguous, and much easier to pronounce and spell than the Latin and Greek epithets currently in use. The mental tyranny of the scholastics would then, at last, be at an end.
No doubt there will be those who will reject the implications of the facts discussed in this section. They will say our taxonomic system is the result of centuries of labor by thousands of naturalists and should not be lightly discounted. Tradition carries weight, they will say. But I simply find puzzling those of my colleagues who think this way and want to go on and on rearranging and patching taxonomies. I see them much as Frost saw his neighbor who insisted on mending walls every spring:
My apple trees will never get across
And eat the cones under his pines, I tell him.
He only says "Good fences make good neighbours."
Spring is the mischief in me, and I wonder
If I could put a notion in his head:
"Why do they make good neighbours? Isn't it
Where there are cows? But here there are no cows.
Before I built a wall I'd ask to know
What I was walling in or walling out,
And whom I was likely to give offence.
Something there is that doesn't love a wall,
That wants it down." I could say "Elves" to him,
But it's not elves exactly, and I'd rather
He said it for himself. I see him there
Bringing a stone grasped firmly by the top
In each hand, like an old-stone savage armed.
He moves in darkness as it seems to me,
Not of woods only and the shade of trees.
He will not go behind his father's saying,
And he likes having thought of it so well
He says again, "Good fences make good neighbours."
NEXT PAGE >>