On the Origins of New Forms of Life
2.8: Triangular Numbers & Section Conclusion
(Continued from the previous page)
|
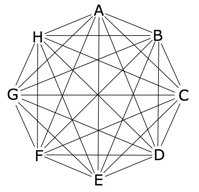
| The pairings between 8 types of organisms (A, B, C, D, E, F, G, and H) are represented by the lines in the diagram above. Note that the number of pairings (28) greatly exceeds the number of organisms being paired. |
Triangular Numbers Show Viable Hybrids are Likely. It is in fact true that attempts at hybridization often end in failure because the hybrids from many crosses turn out to be too infertile or inviable. Nevertheless, the expected number of successes is more than sufficient to implicate hybridization as a major evolutionary force. The reason is straightforward: in the typical group of related forms the number of different types of organisms in the group is generally far exceeded by the number of pairs that can be constructed from those types (see figure at right).
The number of possible combinations from a set of n types is n(n-1)/2. That is, it is a triangular number (a triangular number is the sum of the n integers from 1 to n). For example, one hundred types of organisms can be paired in 100(99)/2 = 4,950 different ways. Thus, even if viable hybrids could be obtained in only one percent of the potential combinations of 100 different types of organisms, there would be 50 successful combinations. This figure does not even take into consideration the additional possibilities introduced by reciprocal crosses, backcrosses, and later-generation crosses.
Therefore the fact that most crosses produce either inviable hybrids, or no hybrids at all, should not be used — as it often is — to discount the significance of hybridization in evolution. Instead of making blanket claims of hybrid inviability and "hybrid breakdown," evolutionary biologists should consider the many well-documented cases of hybrid viability and vigor.
Conclusion. Long ago Darwin (1859: 254) pointed out that hybrids are not always sterile:
Even today, many biologists pay little heed to this fact, although a vast number or reports have since accumulated demonstrating that many hybrids are partially fertile. Indeed, there are probably far more cases of this sort known today than Darwin would ever have anticipated. Section 2 has provided a wide variety of examples demonstrating that the fertility of hybrids varies from cross to cross, and individually, depending on the sex, age, and genetic constitution of the individual in question. It has also shown that hybridization is a widespread natural phenomenon and that many hybrids have even been treated as species or subspecies. A huge number of natural hybrid zones are known.
We have seen that many kinds of hybrids are partially fertile, and that they vary greatly not only in their fertility, but with respect to many other traits. These facts have fundamental implications for biological thought. If many types of natural hybrids have the ability to conceive offspring, the opportunity exists for natural selection to act to increase fertility with each passing hybrid generation. Fertility responds to artificial selection by breeders, much like any other heritable trait that varies from one individual to another. Certainly, fertility varies from one hybrid individual to another (and from one type of cross to another as well). The potential evolutionary significance of hybridization becomes especially apparent if we consider the fact that hybrid vigor is characteristic of a wide variety of crosses.
If stereotypes are left behind and hybrids are viewed as individual organisms with individual characteristics, their varying fertility (and a myriad other variable traits) can be seen in another light — as unshapen clay awaiting the hands of natural selection. NEXT PAGE >>
Most shared on Macroevolution.net:
Human Origins: Are we hybrids?
On the Origins of New Forms of Life
Mammalian Hybrids
Cat-rabbit Hybrids: Fact or fiction?
Famous Biologists
Dog-cow Hybrids
Georges Cuvier: A Biography
Prothero: A Rebuttal
Branches of Biology
Dog-fox Hybrids