On the Origins of New Forms of Life
2.2: Variation in Hybrids
(Continued from the previous page)
 Charles Darwin
Charles Darwin
An organism descended from matings between two or more parental types will mix the characteristics of those types. As Darwin notes in the Origin,
The analysis of a hybrid's traits will therefore often allow the identification of its parents. The traits of parents are expressed in hybrids in two typical ways. They are either intermediate or combined. A general understanding of the nature of trait variation in hybrids is necessary if one is to understand stabilization theory.
Intermediacy. With respect to most traits — size, color, texture, and so on — hybrids are typically intermediate between their parents. For example, Hubbs (1955) evaluated a variety of fish hybrids in the family Catostomidae (suckers). In summarizing his results, he states (p. 5) that
Similarly Grant and Grant (1971b) report that in Arizona natural hybrids between two cholla cactuses (Cylindropuntia spinosior and C. versicolor) show intermediate variation with respect to a variety of morphological traits (e.g., stem joint diameter, tubercle height and width, color of stems, length and number of spines, fruit texture and color).
 Goldeneye
GoldeneyeBucephala clangula
 Hooded Merganser
Hooded MerganserLophodytes cucullatus
In the absence of data from captive crosses, natural hybrids are often identified as hybrids on the basis of their intermediacy. In fact, hybrids are often referred to as "intermediates." Thus, in a detailed study of a natural hybrid between two ducks, the Goldeneye (Bucephala clangula) and the Hooded Merganser (Lophodytes cucullata), Ball (1934) found the hybrid was intermediate between its parents with respect to many characteristics. These included overall length, wing and tail length, width and length of bill, degree of development of head crest, and a variety of other features.
Sturgeon in the Atchafalaya River, a tributary of the upper Missouri River (western U.S.), are hybrids of the pallid (Scaphirhynchus albus) and shovelnose (S. platorhynchus) sturgeons and are also intermediate with respect to a broad range of physical traits and in their genetics. The Flame-rumped Tanager (Ramphocelus flammigerus) has a red rump, and the Lemon-rumped Tanager (R. icteronotus) has a yellow one. These birds hybridize extensively in the Andes. The rumps of their hybrids are intermediate in color, in varying shades of orange (McCarthy 2006).
Intermediacy is seen not only in the physical characters of hybrids but also in their behavior. Evans (1966) reports that hybrids between Sharp-tailed Grouse (Tympanuchus phasianellus) and Greater Prairie Chicken (T. cupido) hold their wings further out during displays than do prairie chickens, but not so far as do Sharp-tailed Grouse. Hybrid hummingbirds often perform dive displays intermediate between their parents' displays.
The territorial displays of hybrid lizards (Anolis aeneus × A. trinitatis) on the island of Trinidad are intermediate too. Koeppl et al. (1978) analyzed the acoustical behavior of hybrids between Richardson's Ground Squirrel (Spermophilus richardsonii) and the Wyoming Ground Squirrel (S. elegans) in southwestern Montana and found their chirps were intermediate between the two typical parental chirps.
Cooper et al. (1998) note that the echolocation call frequency of hybrids between the Eastern and Large-eared horseshoe bats are also intermediate (about 40 kHz, vs. 66 kHz in Rhinolophus megaphyllus and 28 kHz in R. philippinensis). Migration routes of hybrids are often midway between those of their parents (Sibley 1994). Gestation and incubation periods of hybrids are usually intermediate too. Hybrid intermediacy involves the entire organism — any trait can be affected.
 Blue Grouse
Blue GrouseDendragapus obscurus
 Sharp-tailed Grouse
Sharp-tailed GrouseTympanuchus phasianellus
Combined Traits Although intermediacy is the rule, a given type of hybrid may not be intermediate with respect to certain traits, and instead may approach one parental type or another. For example, in birds the plumage pattern of a hybrid's head will often resemble that of one parent, while that of its body will resemble the other. Brooks (1907) collected a wild hybrid between the Blue Grouse (Dendragapus obscurus) and Sharp-tailed Grouse (Tympanuchus phasianellus). He was confused when he first saw this bird because in front it looked like a sharp-tail, but in back it looked like a blue. Kawamura (1950a) found traits of F₁ hybrid frogs between European Common Frog (Rana temporaria) and Japanese Brown Frog (R. japonica) resembled one parent with respect to some traits, but the other with respect to others. Rahman and Uehara (2004) obtained similar results with hybrids between sea urchins in the genus Echinometra.
The vocalizations of hybrids often unite features otherwise found only separately in their parents. Marler and Tenaza (1977) say the song of a female gibbon hybrid (Hylobates lar × H. muelleri) followed this pattern. Jung et al. (1994) describe a junco × sparrow hybrid (Junco hyemalis × Zonotrichia albicollis) which, too, sang a song mixing those of its parents. Likewise, in studying natural hybrids between the Smooth Frog (Geocrinia laevis) and the Victorian Frog (G. victoriana), which occur in southeastern Australia, Littlejohn and Watson (1976a, 1976b) found that hybrid calls combine elements of the parental calls.
Traits are combined even at the molecular level, so that hybrids produce a mixture of proteins, some inherited from one parent, some from the other. Protein subunits of heterogenous origin may join in a hybrid to form a single functional unit. For example, Jiménez-Porras (1967) found that venom from natural hybrids of the Jumping Pit Viper and Picado's Pit Viper (Atropoides nummifer and A. picadoi) is electrophoretically intermediate.
Variation in Later Generations. In a natural setting, pure populations unaffected by hybridization are relatively uniform. F₁ hybrid hybrids between such populations, too, will usually be uniform, even when the two parental forms differ rather markedly. This lack of variation results from each F₁ hybrid hybrid receiving much the same genetic complement from each parental type each time the cross occurs. However, in crosses where later-generation hybrids occur, a wide variety of types can arise because parental traits are variously combined in different individuals. For example, plant breeder Bernice Brilmayer notes that
For this same reason, high levels of morphological variability are a strong indication that a natural population is the product of hybridization (the nature of the genetic variation underlying this morphological variation will be explained in Section 3). Brainerd (1924) describes highly variable later-generation hybrids derived from crosses between various members of the violet genus Viola. He reproduced many of these natural hybrids with artificial crosses. In some of these later-generation hybrids, leaf shape was considerably modified and some of the variants could be stabilized as new types.
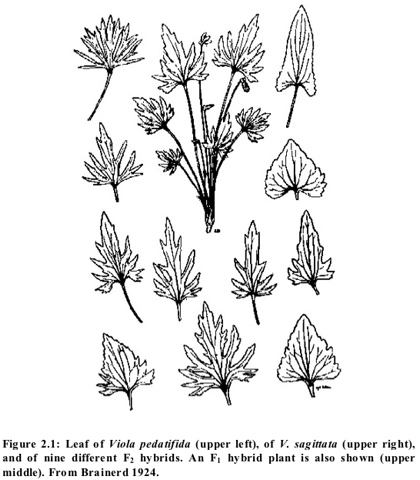
Figure 2.1 shows the leaves of V. pedatifida, V. sagittata, and various F₂ hybrids derived from crossing the two. In reference to a different violet cross giving similar results, Brainerd comments that
Compound hybrids. Compound hybrids are produced by successive matings between more than two different types of organisms. For example, Sandnes (1957) first produced a hybrid between a Lady Amherst's Pheasant and a Golden Pheasant (Chrysolophus amherstiae × C. pictus), and then mated the hybrid with a Common Pheasant (Phasianus colchicus) to produce three-way hybrids. Compound hybrids occur also in the wild. Thus, Harrison and Harrison (1965a) report a presumed natural three-way hybrid of Pintail Duck (Anas acuta) with Mallard (A. platyrhynchos) and Gadwall (A. strepera). In western Mexico, three fish, the headwater livebearer (Poeciliopsis monacha), clearfin livebearer (P. lucida), and chubby livebearer (P. viriosa) produce compound hybrids on an ongoing basis. Camp (1945) argued that Vaccinium corymbosum, the common highbush blueberry of northeastern North America, is the hybrid of four other types that he treated as separate species. Multiple hybrids are common among captive falcons (e.g., Barbary/Gyr × Peregrine/Lanner).
 Geoffroy's marmoset
Geoffroy's marmosetCallithrix geoffroyi
Hybrids with even more complex ancestry are commonly obtained, such as five-way hybrids or six-way hybrids. For example, Coimbra-Filho et al. (1993) produced a six-way hybrid between various marmosets of the genus Callithrix. The ability of plants to produce complex compound hybrids has long been recognized. Already, more than a century ago, Focke (1881) noted that successive intrageneric matings between types treated as separate species can result in five- and six-way hybrids (e.g., in Begonia, Calceolaria, Pelargonium, and Salix). Darwin, too, was aware of compound hybrids:
Most shared on Macroevolution.net:
Human Origins: Are we hybrids?
On the Origins of New Forms of Life
Mammalian Hybrids
Cat-rabbit Hybrids: Fact or fiction?
Famous Biologists
Dog-cow Hybrids
Georges Cuvier: A Biography
Prothero: A Rebuttal
Branches of Biology
Dog-fox Hybrids