On the Origins of New Forms of Life
4.6: Permanent Translocation Heterozygotes
(Continued from the previous page)
|
| Reciprocal Translocation |
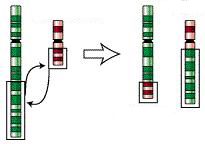
A translocation occurs when a portion of a chromosome is transferred to another location, either on the same chromosome or on some recipient chromosome not belonging to the same chromosome pair (i.e., the same homolog pair) as the donor chromosome. Two chromosomes that belong to the same chromosome pair, that is, that contain the same set of loci in the same order, are called homologous. A reciprocal translocation, the most common kind of translocation, is an exact interchange of chromosomal segments between two chromosomes not belonging to the same chromosome pair (see figure at right).
In a translocation heterozygote two distinct pairs of homologous chromosomes have reciprocally exchanged nonhomologous segments between one member of each pair. As a result each of the affected chromosome pairs contain both homologous and nonhomologous segments. Put another way, each such pair has one translocated chromosome, and one normal (untranslocated) chromosome. More than two chromosome pairs can be altered in this way so that some or all of the chromosome pairs are composed of a translocated and an untranslocated member.
Organisms having chromosomes rearranged in this way are known as permanent translocation heterozygotes. Due to the way reciprocal translocations are processed during meiosis, all the translocated chromosomes pass to one gamete and all the normal chromosomes pass to the other. As a result, only two types of gametes are produced. One has all the translocated chromosomes and the other has all the normal ones (here "translocated" and "normal" are relative terms since in naturally occurring organisms it is usually unknown which of the two types was original). These two sets of chromosomes ("normal" and "translocated") are known as Renner complexes. In well-studied organisms of this type, the various Renner complexes have been assigned formal names.

|
| Oenothera lamarckiana Credit: Frank Vincentz |
{kind=link}
In one of the best-known permanent translocation heterozygotes, the evening primrose Oenothera lamarckiana, the two Renner complexes are called velans and gaudens. So the karyotype of O. lamarckiana is designated as "velans/gaudens." Genetic interchange (crossing-over) between Renner complexes is very rare. So chromosomes in distinct complexes differ genetically in many respects.
Aneuploidization is a common stabilization process among permanent translocation heterozygotes because there is an enhanced tendency to produce aneuploid gametes. The result is the production of numerous trisomic forms. But the most common stabilization process seen in organisms of this type occurs when forms with distinct Renner complexes hybridize to produce new forms. For example, hybridization of O. lamarckiana with another evening primrose, O. strigosa, which has the Renner complexes deprimens and strigens, yields four different types,
- velans/deprimens,
- velans/strigens,
- gaudens/deprimens,
- gaudens/strigens
Some hybrids produced by such recombinations of Renner complexes are reproductively stable and others are not. By this means new stable types can be produced in a single generation. O. lamarckiana itself arose by such a process via hybridization between O. biennis and O. hookeri. This event occurred within the last two or three centuries in Europe where the two parental forms, both native to North America, were introduced. O. lamarckiana spread across Europe and later to North America. In fact, most of the 18 forms of European Oenothera that Renner (1942) treated as species arose in this way. NEXT PAGE >>