
The Hybrid Hypothesis
6: Additional Evidence
|
The first distinguishing characteristic of thinking then is facing the facts — inquiry, minute and extensive scrutinizing, observation.
—John Dewey Reconstruction in Philosophy |
Continued from previous section
Hi. I'm stunned, amazed, and converted.
Gathered from diverse sources, the traits included in this section have not previously appeared on a single list. Any one of them, encountered in isolation, could be shrugged off as insignificant or coincidental. Taken together, though, and in the wake of information considered in previous sections, their implications become more intriguing. Why should the pig, usually considered no more closely related to us than a dog, have a close affinity to humans with regard to a broad spectrum of traits that distinguish us from nonhuman primates? Why should this animal be a stellar performer in the fields of human biology and medicine? As Britt points out, "Pigs' physiological similarities to man—surpassed only by the primates'—make the animals invaluable to medical science" (Brit 1978, p. 406). Many other authors echo this opinion (see: Tumbleson 1985). Why the pig? The situation is even more puzzling if we consider the many cases that we have already examined where pigs actually exceed nonhuman primates in similarity to human beings.
SURFACE LIPIDS
The principal structural components of living cells are proteins, carbohydrates, and lipids. Lipids are fats and fat-like substances, only sparingly soluble in water. Nicolaides et al (1968) compared the skin lipids of human beings with those of eighteen different types of mammals (including a pig, a baboon, a chimpanzee, as well as others: horse, cow, sheep, goat, dog, cat, rat, mouse, hamster, rabbit, guinea pig, chicken, duck, goose, and turkey). They note (Nicolaides et al. 1968, p. 88) that human surface lipids are rich in triglycerides and free fatty acids, but that "none of the other animals [in the survey] had significant amounts of triglycerides or free fatty acids in their skin surface lipid except the pig … Additionally, pig surface lipid showed the same lipid classes as those obtained from the nonpolar epidermal lipid samples of different [i.e., various] human sources, namely, sterol esters, triglycerides, free fatty acids, and free sterols for sole epidermis."
MELANOCYTES
Melanocytes are found in the dermis of panins (see green box, right), and most other nonhuman primates(Montagna 1972, p. 115b), but not in the dermis of human (Cichorek et al. 2013; Montagna 1962, p. 77) or pig (Montagna 1964, p. 13), where these pigment cells are limited to the epidermis. Skin expert William Montagna (1962, p. 77) states that "In man, melanocytes are found consistently in the dermis only during fetal life, and in some nevi. Many other primates, however, have rich populations of them." In addition, according to Montagna (1969, p. 334; see also Montagna 1962, p. 243; 1964, p. 13; 1971, p. 587; Montagna and Yun 1963, p. 194), humans and pigs (but no other known mammal) have melanocytes in the hair follicle matrix.
ACNE
In discussing various human diseases not seen in chimpanzees, Varki (2000, p. 1068a) states that “Acne vulgaris, a common skin affliction of human teenagers also appears to be uncommon in the adolescent chimpanzee.” Nevertheless, Ganderup (2012, Table 21.4) lists it among the various human conditions that can be studied in swine.
It has been recognized for several decades that exposure to language plays an essential role in the development of intelligence. Deaf children who are not exposed either to spoken or sign language at a sufficiently early age develop significant, permanent mental handicaps. So it may well be that the presence of vocal cords in humans was essential to the initial emergence of human intelligence. Thus, it's easy to see why humans (if they really are pig-ape hybrids) would have taken an upward leap in intelligence after inheriting a panin-like brain made heterotic by a pig-like cooling system (as discussed in the previous section), plus a pig-like vocal apparatus (in particular, vocal cords). After all, even human beings with brains that are structurally normal, never achieve normal cognition without language. So an imaginary panin with a fully human brain, but who lacked vocal cords, would not be expected to exhibit a fully human intelligence.
The Island of Dr. Moreau
MELANOMA
Malignant melanoma is a deadly form of cancer that initially affects cutaneous melanocytes and rapidly spreads to other parts of the body. Melanoma in swine closely parallels melanoma in human beings. I have, however, been unable to locate any observation of melanoma in a primate; apparently none has been recorded. Schultz (1956, p. 969; see also O'Gara and Adamson 1972) notes that all types of cancer are rare in nonhuman primates (detailed information about the rarity of cancer in nonhuman primates >>).
VOCAL CORDS
In The Descent of Man, Darwin (1871) asserted that "the fact of the higher apes not using their vocal organs no doubt depends on their intelligence not having been sufficiently advanced." But later anatomical studies have shown that the panin vocal apparatus is not suitably constructed to produce human speech. Thus, after dissecting the larynx of a chimpanzee, Kelemen (1948, p. 254) reached the conclusion that "even with the help of the high grade of intelligence of this animal every attempt to make him utter human voice and speech must fail on the basis of anatomy. An imaginary being equipped with a human brain and the larynx of a chimpanzee could not produce any other phonetic effect than this animal actually does." Although panins can be taught to communicate visually using symbols and gestures, efforts to teach them to speak have failed miserably.
A key difference between humans and panins is found in the structure of the vocal cords. A horizontal slit (ventricle) marks each of the two interior sides of the human larynx. The upper borders of these ventricles are defined by the superior vocal cords, also known as the false vocal cords because they are not concerned with the production of sound (Gray 1977, p. 960), which contain the ventricular ligaments (Gray 1959, p. 1182). On their lower edges, the ventricles are bounded by the inferior vocal cords, which, in humans, contain strong bands of yellow elastic tissue (vocal ligaments) that can be placed under tension to produce vibrations (voice) as air escapes from the lungs (Gray 1977, pp. 960, 1182).
In contrast to the human situation, Sonntag (1923, p. 398) found that the inferior vocal cords of the chimpanzee "are soft and flaccid, consisting almost entirely of mucous membrane, and a little soft tissue. The false cords are likewise soft, and between them are well-marked ventricles." Because they are soft and contain no vocal ligaments, panin vocal cords cannot be tensed, and so, cannot produce human-like vocal tones. They are vocal in name only. Pigs, however, do have a ligament on both sides of each ventricle (Getty 1975, p. 122), in positions corresponding to the human vocal and vestibular ligaments (Sobotta 1983, figs. 317, 318). So although Darwin is no doubt correct in suggesting that human intelligence plays a critical role in the production of speech, a second essential ingredient of human phonation is found in pigs, but not in panins.
LARYNGEAL SACS
The great apes have laryngeal air sacs, large, irregularly shaped, air-filled organs opening into the larynx through the ventricles and spreading outward from the larynx through much of the neck and chest (Nishimura et al. 2007; Raven 1950, p. 23 & figs. 2, 24, 25; Schultz 1968, p. 141; Sonntag 1923, pp. 333, 398). The function of these organs is unknown. They are not present in pigs and human beings.
 Pig stomach (Sus scrofa)
Pig stomach (Sus scrofa)
STOMACH
The stomach of the human fetus passes through a stage during which it has a diverticulum ventriculi, i.e., a small side cavity in addition to the main stomach chamber (Figure 9.1). This blind pouch is commonly found as a transient differentiation at the upper end of the stomach in human fetuses. This feature does not recapitulate anything seen in the stomachs of nonhuman primates. The stomach is simple (single chambered) in all primates except the langurs and the guerezas, which have stomachs that are highly aberrant by primate standards, having no resemblance to the human organ (Duckworth 1915, p. 149, Fig. 94; Sonntag 1924, p. 9). A simple stomach has only a single chamber. The stomach of the domestic pig is shaped like that of a human being, but it does have a small blind pouch identical to that seen in human fetuses (Shively 1984, p. 495). Why should the human fetus exhibit the same pouch in the same position if human beings are related to apes alone? These pouches occur, too, in human adults, but at a low rate of incidence, and seventy-five percent are located 2 cm below the esophagastric junction and 3 cm from the lesser curve (Farhan et al. 2012), in other words, in the position shown for pigs in the illustration (see also Hirsch 1903). It is easier to account for these facts under the hypothesis that humans are pig-ape hybrids. At any given stage of development, a hybrid combines traits derived from each of its parents.
VALVULAE CONNIVENTES
In nonhuman primates the small intestine contains no circular valves (valvulae conniventes). Also called the “valves of Kerckring”, these circular membranous folds are found on the interior surface of the small intestine of humans and certain animals. They are so large that they may be felt through the wall of the intestine. Nelsen (1953, p. 699) notes the presence of "the valves of Kerckring in the human and pig small intestine" (ibid. see also p. 629, fig. 298b). These structures are thought to enhance the absorption of nutrients by increasing the internal surface area of the intestine.
MESENTERY
In human beings the superior mesenteric artery supplies blood to the small intestine and colon. Connecting it with the small intestine are the intestinal arteries, which form an arcade (a series of interconnected arches); this arterial arcade is connected to the intestine by numerous, small straight vessels (Gray 1959, pp. 678-679 & Fig. 577). No similar configuration is found in panins (Sonntag 1924, p. 241). The pig, however, does have human-like arterial arcades connecting the superior mesenteric with the small intestine (Getty 1975, p. 1332; Nickel et al. 1973, Fig. 145).
HEART
"In human hearts placed on their apices, both the auricles lie on nearly the same level. In chimpanzees, as in the rest of the Pongidae, the left auricular appendage lies distinctly lower" (Frick 1960, p. 299). Sisson and Grossman (1953, p. 754, fig. 605) show that the auricles of a pig heart are approximately level with each other.
Heart attack is, of course, a very common cause of death in human beings (as it is in pigs when they are allowed to get old instead of being slaughtered young (Spörri 1954). Sporri mentions “the marked predisposition of the pig to death from heart failure.” While panins and other primates do experience various forms of heart disease (Ruch 1959), they do not have heart attacks like those of human beings. This fact puzzled Varki et al. (2009), as evidenced by their comments:
When the tricuspid valve of a human heart is diseased, currently accepted medical practice specifies its replacement with a pig tricuspid valve—not that of a dog or a baboon (Brit 1978, p. 406). In fact, according to Osther et al (1991), "Porcine products have been used extensively in many human therapeutic replacement regimens. Their use is primarily due to genetic similarity of humans and pigs at the amino acid level, which results in a high acceptance of grafted prosthetics and excellent tolerance of repeatedly administered biologicals." For example, pig skin is used in treating human burn patients (Aronoff et al. 1976; Brit 1978, p. 406). Human diabetics are given pig insulin.
ATHEROSCLEROSIS
In biomedical research the pig has proven to be the model of choice for investigating human atherosclerosis, another commonly fatal disease in humans. Pigs are much preferable in this respect to nonhuman primates (Pond and Houpt 1978, p. 32). In fact, they develop human-like atherosclerosis at a much earlier age than do humans themselves, and the condition occurs spontaneously, that is, it develops even in pigs eating a low-fat diet. In a study of “aged” hogs (i.e., sows aged 8-12 years of age), Luginbühl and Jones (1965, pp. 19-20) state that
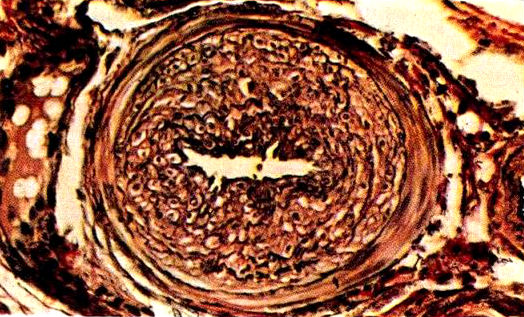 Atherosclerosis in a swine: cross-section of a nearly occluded coronary artery (source: Luginbühl and Jones 1965, Fig 1-9).
Atherosclerosis in a swine: cross-section of a nearly occluded coronary artery (source: Luginbühl and Jones 1965, Fig 1-9).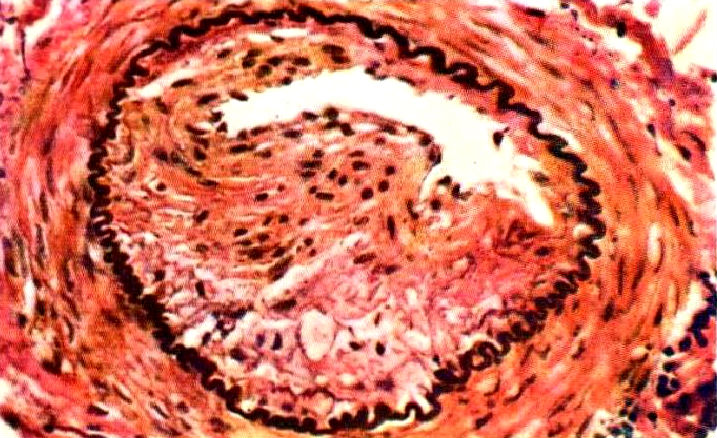 Atherosclerosis in a swine: cross-section of a nearly occluded cerebral artery (source: Luginbühl and Jones 1965, Fig 1-18).
Atherosclerosis in a swine: cross-section of a nearly occluded cerebral artery (source: Luginbühl and Jones 1965, Fig 1-18).
Zubige (1965, p. 37, citing Florey 1963, Jennings et al. 1961, Rowsell et al. 1958, and Skold and Getty 1961) says that
“In apes, atherosclerotic lesions are absent or minimal despite reported high cholesterol levels” (Munson and Montali 1990, p. 103; see also Benirschke and Adams 1980). Clarkson (1965, p. 211) goes so far as to claim that coronary atherosclerosis in chimpanzees is unknown. And Strong et. al. (1968, p. 883) state that “It should be emphasized that
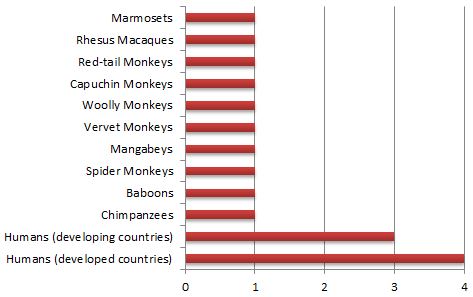 Atherosclerosis severity in primates on a scale of 1 to 4 (source: Strong et al. 1968, Table 1).
Atherosclerosis severity in primates on a scale of 1 to 4 (source: Strong et al. 1968, Table 1).
KIDNEYS
In terms of gross morphology, the human kidney bears little resemblance to the kidneys of other primates—for example, Elftman and Atkinson (1950, p. 201) speak of "the fundamental contrast between the human kidney and that of primates in general"—and is certainly much more similar to that of a pig than to that of a panin. As can be seen in the figure below, pig kidneys have the same bean-shaped cortex seen in humans, while a chimpanzee's is almost triangular. The interior cavity of this organ is large in humans and pigs, but small in chimpanzees. Note especially the numerous inward projections (pyramids) in the human and pig kidneys. In the ape kidney, the projection is single, centrally placed, and quite flat. This latter type of configuration is termed unipyramidal. The configuration in human beings and pigs is termed multipyramidal. In this respect, human kidney structure contrasts sharply not only with that of panins, but also with that of all other primates (Hill 1966, p. 44).
 |  |  |
| Chimpanzee kidney | Human kidney | Pig kidney |
Pig kidneys are similar to those of human beings not just in being multipyramidal. The similarity extends to fine details such as regional distribution of pyramid type (compound vs. simple), renal lymphatics, and distribution of pacemaker cells (Ransley and Risdon 1975; Terris 1985). The pig is the only creature other than human beings known to have this type of kidney, that is, a multipyramidal with a smooth, bean-shaped, undivided cortex (Terris 1985). After personally dissecting 117 kidneys, representing 67 animals of various primate genera (including Pan), Straus (1934, pp. 97-98) emphasized the sharp distinction between the human kidney and those of other primates:
Elsewhere in the same paper, Straus (1934, p. 96) notes that he never observed more than two pyramids in any spider monkey kidney. Certainly, then, humans are peculiarly isolated from other primates; Homo averages ten pyramids per kidney, as does the domestic pig (Sisson and Grossman 1953, p. 579). But renal pyramid count seems not, as Straus claims, to be devoid of taxonomic value: It is suggestive of a link between human and pig. Straus’ attitude toward this trait exemplifies a more general problem: Biologists tend to dismiss data that contradict accepted notions concerning the nature of phylogenetic relationships.
BACULUM
"In all primates except Tarsius [tarsiers], Lagothrix [South American wooly monkey] and man, the distal part of the penis is strengthened by a rod-like bone (baculum)" (Hill 1966, p. 44; 1972, pp. 113, 115). Dixson (1981, p. 56) also lists spider monkey (Ateles) as an exceptional nonhuman primate possessing a baculum. Thus, human males lack a structure common to virtually all their primate kin, including all apes (Martin and Gould 1981, p. 143). This finding is consistent with the idea that humans are pig-ape hybrids: the pig lacks a baculum (Sisson and Grossman 1953, p. 602).
PENIS
It is sometimes claimed in the literature that Homo sapiens has the largest penis of any primate (252.5; 385.5,10,58), but photographic evidence clearly demonstrates that the erect chimpanzee penis is comparable in size to a human’s (585.3,127). The obvious feature differentiating the penis of Homo from that of Pan is the presence of a glans penis. The panin glans clitoridis is almost identical in form to the glans penis in Homo, but a male panin penis has no expanded glans (36.2; 140.5,429,Fig. 3h; 235.7,115; 252.5; 490.3,Plate 4; 533.8,270; 585.3,127). Gorilla males also have a glans similar in shape to that of chimp females (140.5). This glans structure may have been transferred between the sexes in humans and gorillas due to the chromosome rearrangements that would have occurred in later-generation pig-panin hybrids. Banding studies lend some support to this idea because both gorillas and humans have a larger Y than do panins (269.5; 317.8; 613.5) and, in both cases, the Y shows brilliant quinacrine fluorescence not seen in the panin Y, or in the Y of any other primate (348.6; 429.93; 541.6). The panin X, however, does fluoresce brilliantly (as do, also, those of humans and gorillas). It could well be, then, that a piece of the brilliant panin X has been transferred, during the process of chromosomal rearrangement characteristic of recombinational stabilization, to the Y in Homo and Gorilla and that the traits common to the human penis and panin clitoris are specified by genes present in this transferred chunk of DNA (subsequent discussion will eventually explain why it seems likely that gorillas are also of hybrid origin).
HYMEN
"The presence of the hymen is always cited as a human peculiarity, and several authorities have definitely asserted its absence in the anthropoids. Sonntag has declared that it is absent in all apes" (Jones 1929, pp. 323-324). Duckworth (1915, p. 209) says the hymen is present in "the hominidae alone among the primates." In particular, a hymen is absent in the chimpanzee (533.6,420). At the time Duckworth was writing, the family Hominidae included humans alone, whereas it now generally understood to encompass, also, the great apes. Nickel et al. (1973, pp. 376-377), note of the pig that "at the junction [of the vagina] with the vestibule, in the virginal animal, is an annular fold 1-3 mm. high, homologous to the hymen."
SCROTUM AND TESTICLES
Hill (1966, p. 44) asserts that the scrotum is sessile in chimpanzees and gorillas (i.e., that it does not hang away from the body). This is accurate with regard to gorillas, but a photograph by van Lawick (1986) of a mature chimpanzee male demonstrates that the chimpanzee scrotum is pendulous. Moreover, Roberts (p. 880) states that "a pendulous scrotum and penis are characteristic of primates." A scrotum that hangs down, then, is not a human distinction, as some authors have claimed. With respect to the size of their testicles, both panins and pigs greatly exceed human males (Schultz 1938). The lack of human intermediacy can here be ascribed to hybridity. Hybrids often have underdeveloped testicles.
PROSTATE
The prostate gland is a firm, partly muscular, partly glandular organ situated at the base of the male urethra just below the bladder. It secretes a sticky, alkaline fluid which is a major constituent of semen. In primates, "the prostate is generally bi-lobed, a traverse groove dividing it into a cranial and caudal lobe, but only in Homo and Ptilocercus‡ does it completely encircle the urethra. In other primates it leaves the ventral wall of the passage uncovered" (Hill 1972, p. 115; see also Mijsberg 1923; Raven 1950, p. 88a). In boars, however, it does form a complete circle (Getty 1975, p. 1300; Sisson and Grossman 1953, p. 602), just as it does in men.
BULBOURETHRAL GLANDS
Also known as Cowper's glands, the bulbourethral glands are found only in males. In humans they are about the size of a pea and are located on either side of the prostate gland. Each has a duct about an inch long, terminating in the urethra. During copulation, the pre-ejaculatory fluid secreted by these glands serves to clear the urethra of urine and to lubricate the penis and vagina. Bulbourethral glands "have never been reported in anthropomorphs [i.e., apes] in spite of diligent search" (Hill 1966, p. 45). Nevertheless, large bulbourethral glands are found in boars (Getty 1975, p. 1300).
COITUS AND SEXUAL CLIMAX
Morris (1967, p. 63) notes that in nonhuman primates "pre-copulatory [behavior] patterns are brief…Copulation itself is very brief." He cites the baboon, whose time from intromission to ejaculation is only "7-8 seconds," with no more than 15 strokes. The duration of coitus in anthropoid apes is scarcely longer. The pygmy chimpanzee (bonobo) averages 24.7 strokes per intromission (Savage-Rumbaugh and Wilkerson 1978, p. 336) and the common chimpanzee only 11.3 strokes (ibid. citing Tutin and McGrew 1973). This figure is about 36 in gorillas (Nadler 1976). Dixson (1981, p. 149) says chimpanzees ejaculate five to ten seconds after intromission. The human male, then, who takes several minutes or more to reach climax (Masters and Johnson 1966), stands in marked contrast to the apes.
In swine, however, coitus lasts 20 to 30 minutes. In itself, the ejaculation of a boar can continue nearly ten minutes (Hughes and Varley 1980, p. 200; Sack 1982), and the volume of the ejaculate can be as much as a liter, although the average is closer to 200 ml (Sack 1982). Domestic boars have been observed to ejaculate as many as 25 times in a single day (Bustad and Horstman 1985, p. 12; Hedgepeth 1978).
Given the lengthy attentions lavished upon the sow by the boar, it is, perhaps, not surprising that "the female [pig] exhibits an 'orgasm-like' response" (Hafez 1968, p. 162). Sexual climax has not been observed in nonhuman primate females. Morris (1967, p. 79) says that "female orgasm in our species is unique amongst primates." "If there is anything" he says (ibid. p. 63), "that could be called an orgasm [in nonhuman primates], it is a trivial response when compared with that of the female of our own species."64 Others affirm this fact.† If we are related only to primates, then it is odd that female orgasm occurs in humans but not in other primates.
AFFECTION AND GRIEF
Pigs are also very affectionate animals, capable of forming strong attachments. Following the loss of a close associate, many pigs exhibit a grief-like syndrome "causing the animal to reject food, be depressed and frustrated, angry and to search almost frantically for the missing animal or person. In some instances, the animal may die" (Bustad and Horstman 1985, p. 13). Pigs enjoy close physical contact and like to snuggle while sleeping.
In contrast, excepting mother and infant, chimpanzees sleep alone (Goodall 1983). Famous chimpanzee observer Jane Goodall (ibid., p. 194) writes, "Chimpanzees usually show a lack of consideration for each other's feelings which in some ways may represent the deepest gulf between them and us. For the male and female chimpanzee there can be no exquisite awareness of each other's body—let alone each other's mind. The most the female can expect of her suitor is a brief display, a sexual contact lasting at most half a minute, and, sometimes, a session of social grooming afterward. Not for them the romance, the mystery, the boundless joys of human love."
UTERUS FORM
In a bicornuate uterus, the main body of the organ is divided into two long chambers ("horns"). A bicornuate uterus is the normal configuration found in swine and most other domestic animals (Nickel et al. 1973, p. 376, Fig. 543; Sack 1982), as it is, also, in lemurs and lorises (Hill 1972, p. 116). All monkeys and apes have a simple, undivided uterus, which is also the condition found in most humans (ibid.; Banks 1974, p. 250). But according to Luckett (Luckett 1974, p. 209) "a bicornuate uterus occasionally occurs in the human, and pregnancy may ensue and proceed to term under these conditions."
MENSTRUAL CYCLE
The chimpanzee menstrual cycle lasts 36-37 days;72 the human, an average of about 28; and the porcine, 21.73 The human cycle is intermediate (as is typically the case with hybrids of other kinds).
SEXUAL SWELLINGS
Panin females (and females of many other kinds of primates) develop large sexual swellings. These structures shrink away as the female goes "out of season." In panins, they are pink, larger than in most other primates, and have a stimulating effect upon the male.74 No similar swelling of the female genitals occurs in human beings or pigs. In fact, a breeder has to be something of an expert to be certain whether a sow is ovulating or not.
LABIA
Flanking the entrance to the urino-sexual opening of human females are two sets of lips, differing in structure and size (Gray 1977, pp. 1025-1026). The external lips, the labia majora, are thicker and coated with ordinary skin. The labia minora lie inside and are more delicate in structure and are coated with epithelial tissue. The typical nonhuman primate has labia minora, but not labia majora (Kraemer and Vera Cruz, p. 843). In particular, "the chimpanzee's vulva never, at any age, attains the typically human form of a simple linear pudendal cleft, guarded by two parallel cutaneous folds (labia majora) which hides from view, in the undisturbed state, the labia minora, the clitoris and its praeputial appendages and all deeper structures … true labia majora, lying more or less parallel to the median cleft, are found only in foetal life, as in monkeys" (Hill 1951; see also Sonntag 1923, p. 420). A sow's vulva exhibits the opposite configuration, having labia majora only (Nickel et. al 1960, p. 377), and so is quite similar in external appearance (Smith et al. 1990, fig. 250) to the corresponding human structure. The human configuration, then, can be accounted for by assuming labia minora are derived from panins and labia majora, from pigs.
ISCHIAL CALLOSITIES
Ischial callosities are large, thick, hairless patches of skin seen on the rumps of nonhuman primates. When these animals sit, their callosities provide a tough cushion between pelvis and ground. "Humans are unique among catarrhines [i.e., humans, apes, and Old World monkeys] in their total lack of ischial callosity development" (Schwartz 1987; see also Schultz 1936, p. 266; 1968, pp. 139-140). Ischial callosities are not found on a pig's rump either.
BREASTS
| Jones (259.8,324) says "One other well-known peculiarity of man … is the fact that the human nipples are situated considerably lower on the chest than they are in any other known primate." See also (495.06,281 & 445,Table 6). Schultz (495.65,140) notes that, among primates, "in man alone do they [i.e., the nipples] migrate caudally [downward] during growth." Pigs have nipples on their bellies (405.9). | ||||
|
| ||||
|
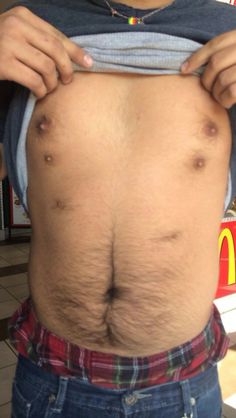
Supernumerary nipples
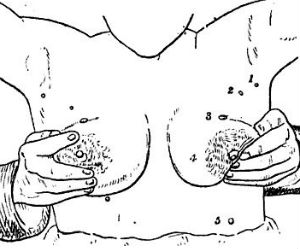
Supernumerary (extra) nipples are fairly common in human beings (Brown and Schwartz 2002). In women, these extra nipples produce milk during lactation. The prevalence of supernumerary nipples varies, depending on the population, but in certain ethnic groups, such as Arabs and Japanese, they are present in one person in twenty (see Brown and Schwartz for prevalences in various human populations). They usually arise within the embryonic milk lines, that is, they occur along lines corresponding the the lines of nipples present in various animals such dogs, cats, and pigs. I have been unable to find any reported case of a supernumerary nipple in a non-human primate (in these animals the normal number of nipples is two). If anyone knows of such a case, please forward the relevant citation to this website. In humans supernumerary nipples are most commonly single. But cases are known in which as many as ten have been present, for example, the woman pictured in the illustration below.
|
"Except in Homo, mammary glands, during the quiescent phase, are poorly represented [in nonhuman primates], and even in the gravid phase and during lactation they do not form external swellings, enlarging instead diffusely subcutaneously. The only external manifestation is an enlargement of the nipples, which elongate and become pendulous" (Hill80). On the other hand, enlarged breasts are characteristic of mature human females. During lactation, sows' breasts also enlarge and are similar in form to those of human females.81 In this respect, female pigs are more similar to human females than are nonhuman primates. Nevertheless, nonlactating breast enlargement is a characteristic that distinguishes Homo even from Sus.
This discrepancy, perhaps, can be accounted by considering how certain traits are combined in human beings. A typical mammary gland is divided into chambers (milk sinuses). Each sinus is connected to the teat by a separate milk duct. The number of milk ducts, and, hence, the number of milk sinuses, in a single human mammary gland ranges from 15 to 22.82 In the chimpanzee, the range is narrower, from 20 to 22.83 In contrast, pig mammae have a relatively simple structure, being divided into only two or three milk sinuses.84 As noted in the section on human skin, a thick layer of subcutaneous fat (panniculus adiposus) is characteristic of pigs and human beings, but not of panins. In the human breast this fat is interspersed between the milk sinuses, as it is in pigs. The volume of the interspersed fat is much greater in human beings because numerous milk sinuses imply numerous intervening spaces for fat storage. Fat is the major constituent of the nonlactating human breast. If each of the numerous sinuses of the chimpanzee mammary were ensheathed in fat like human sinuses, then the mammary region might be as prominent in chimpanzees as in humans. The novel combination of a panniculus adiposus with a complex sinus system may thus be responsible for nonlactating breast enlargement in human females.
ALCOHOLISM
Although Cheeta, Tarzan's chimpanzee sidekick, often drank beer when he wasn’t making movies,85 nonhuman primates are not heavy drinkers—even when they get the opportunity. On the other hand, according to Tumbleson, "the pig is the ideal model for human alcoholism because it is the only mammal other than man that will drink enough ethanol to be classified as an alcoholic."86 One of Tumbleson's pigs drank the equivalent of four quarts of 86-proof vodka every day for a week. The average pig in Tumbleson's herd was a more modest consumer, taking in only a quart a day. Moreover, pigs drink alcohol, even distilled alcohol, voluntarily, if it is available. With continued use they become dependent on alcohol, exhibiting withdrawal symptoms similar to those seen in human alcoholics.87
TEARS
When upset, chimpanzees whine and moan, but they never shed tears.88 Since tears lack commercial significance, little attention has been directed towards this particular physiologic response in pigs, but according to Dexter et al., one of the withdrawal symptoms seen in distressed alcoholic pigs is lacrimation (i.e., the secretion of tears).89
NOCTURNAL ACTIVITY
With the exception of the South American night-monkey (Aotus), monkeys and apes are not active at night.90 They retire with the setting of the sun. This inflexible behavior pattern is by no means characteristic of Homo sapiens. By choice, pigs are active during the day, but when they are hunted they follow a nocturnal schedule.91
TERRESTRIALISM
The typical primate, with the two notable exceptions of baboons and gorillas, spends much, or even most, of its time in an arboreal setting. Such, of course, is not the case with humans and pigs.
SWIMMING AND BATHING
In general, primates have a greater fear of water than the members of perhaps any other mammalian order. Until very recently, ape experts generally believed that panins could not be trained to swim. Thus, Vernon Reynolds in his book, The Apes observes that
But in the July 2013 issue of American Journal of Physical Anthropology ("Swimming and diving behavior in apes (Pan troglodytes and Pongo pygmaeus): First documented report") Bender and Bender document that chimpanzees and orangutans actually can be trained to swim. So the ability to swim is not, as has long been thought, a characteristic that clearly distinguishes humans from chimpanzees (to see videos of a chimpanzee and an orangutan swimming, click here). It still appears safe to say, though, that non-human primates generally have a strong aversion to water and do not learn to swim unless they are raised by humans and taught to do so.
In contrast, the domestic pig, "like many of its wild relatives, is very fond of water. Pigs can obtain food underwater, are fond of fish and are very capable swimmers" (Bustad and Book93). The 1984 recipient of the American Humane Association Stillman Award for animal heroism was a pet pig named Priscilla who earned the prize by saving a drowning 11-year-old boy. Pigs are even capable of catching fish underwater.94 Diving pigs, for example, regularly perform for the public at Aquarena Springs in Texas. A pig is at least as good a swimmer as most human beings.
Given access to water, pigs not only swim, but also use it to keep themselves clean. The actual behavior of pigs defies the stereotype that casts them in the role of filthy mud-wallowers. According to swine experts L. K. Bustad and V. G. Horstman, "Under normal conditions, pigs are instinctively neat and fastidiously clean. They much prefer a clean dry bed and are exceedingly fond of comfort and warmth."95 Pigs confined to a muddy pen are forced to be dirty, just as humans would be under similar circumstances. When given the option, pigs are actually rather tidy. For their pigs, Bustad and Book "provided outside runs with a cold water misty spray. Our swine would select a remote area in an outside run for their defecation and urination, and appeared very neat and clean under these conditions."96 Swine are easily housebroken.97
In contrast, apes do not bathe even when given the opportunity. Their bodies constantly emit a powerful, reeking odor. This disregard of personal hygiene is also reflected in a rather nonchalant attitude toward bodily wastes—despite their much vaunted intelligence, they are difficult to house train. "In their habits of excretion, the apes have not developed specific patterns of behavior. They have no defecating areas, and no set times for defecation. This is typical of primates. Man learns the place to defecate; but wild apes do not, and apes raised in captivity mostly do not either" (Reynolds98).
OMNIVOROUS DIET
At one time it was believed that Homo sapiens was the only meat-eating primate. In recent years, however, researchers have shown that meat and insects are important in the diets of many primates.99 Chimpanzees not only eat meat, but hunt together in cooperative groups.100 There are even well-documented cases of chimpanzees preying on human infants,101 and elsewhere on this website there is even a video showing a chimpanzee troop hunting down a weakened chimpanzee and eating him (watch the video >>). But chimps more typically attack small animals such as monkeys and the young of ungulates.102 It may be, however, that chimpanzees do not digest meat as well as human beings. Goodall studied the diet of wild chimpanzees, using a method called dung-swirling (swirling of dung in water to separate its components for analysis). She states that "it is amazing how much of a chimpanzee's food seems to pass through the digestive tract only partially digested," and notes in particular that "dung-swirling was an excellent method of determining how often members of our group ate insects and meat."103 I assume that the method was a good one for insects and meat because foods of this type were not well-digested. If this assumption is correct, then chimpanzees are not as omnivorous as humans. Certainly, meat cannot pass through the human digestive tract and remain in recognizable form. Nor would it pass through a pig unscathed.
The pig is the archetypical omnivore. Walker104 notes that foods eaten by wild pigs include fungi, roots, tubers, bulbs, green vegetation, grains, nuts, cultivated crops, and animals, both vertebrate and invertebrate—a list that corresponds rather closely with the items normally found on the human menu. "In its nutrient requirements," say Pond and Houpt, "the pig resembles humans in more ways than any other mammalian species."105 One of the main attractions of pigs as domestic animals has been their willingness to eat all of the leftovers from the farmer's table. In fact, the porcine digestive tract is so efficient that it can even extract nutrients from human feces. Thus, if human beings really are more omnivorous than panins, a close relationship to pigs would explain why.
APPENDIX A: NOMENCLATURE OF THE MULTIPYRAMIDAL KIDNEY
Straus (1934, pp. 97-98) found only one renal pyramid in nonhuman primates. This finding is supported by von Bischoff (1877), Bolau (1876), Chapman (1881, p. 168), Ehlers (1881), Elftman and Atkinson (1950, pp. 200-201), Gagnon (1970), Hill (1966, p. 44), Sperber (1944), Symington (1891), Tisher (1970), and Weber (1904). Elftman and Atkinson note that Gerhardt (194.3) described a gorilla kidney as "multipyramidal" but showed unipyramidal kidneys in his illustration (Plate III, 13a and 13b), and therefore reach the conclusion that "it seems highly probable that the kidneys described by Gerhardt bore a closer resemblance to those of man in verbal description than in actual morphology, due to the confusion of nomenclature reviewed by Straus [i. e., Straus 1934]" (Elftman and Atkinson, 1950, p. 201).
This confusion was over the terms "pyramid" and "sub-pyramid." Straus points out that certain writers (Deniker 1885, Sonntag 1923; 1924, Jones 1929) used the term pyramid (or papilla) to describe the structures called "sub-pyramids" by most other writers. He (Straus 1934, p. 97) notes that this is made clear by Deniker's illustrations, as well as by Jones' and by Sonntag's use of the term "fused pyramid." Because sub-pyramids are subdivisions of pyramids, they will obviously always be more numerous than pyramids themselves. When Sonntag (1923, p. 399) says that "Symington [1891] found the pyramids fused to form one papilla," or that in the chimpanzee "The blunt apices of the pyramids do not project much, and they are not embraced by large calyces," it becomes obvious to anyone familiar with the actual renal anatomy that Sonntag is referring not to pyramids, but to subpyramids. I include Straus' discussion of the nomenclature problem:
Hou-Jensen [1930], however, has convincingly demonstrated that the above concept of renal pyramid is neither correct (historically) nor satisfactory. According to him, the pyramid is a division of the medulla, completely isolated by inward cortical projections (true renal columns or septa interpyramidalia) and ending in its individual and isolated papilla. Hence the number of pyramids and papillae exactly coincide. The pyramid is usually divided into two or more secondary pyramids by less extensive inward projections of the cortex (false renal columns or septa pyramidis).
The difference between the above two descriptions of the human kidney is really but one of definition. Yet that of Hou-Jensen is more satisfactory in that it recognizes and classifies minor subdivisions of the medulla. Thus his definitions of pyramid, etc., are more applicable to comparative renal study, and have therefore been adopted by the present writer.
It is this standard definition of "pyramid" that is employed in the present work to compare humans, apes and pigs.
CITATIONS:
1. (87.6,406)
2. (577.6)
3. (410.5)
4. (410.5,88)
5. (360.8,115b)
6. (103.45; 360.1,77)
7. (360.3,13)
8. (360.5,334). See also (360.1,243; 360.3,13; 360.7,587; 365.5,194).
9. (495.44,969). See also (413.2).
10. (135.6)
11. (269.3,254)
12. (220.1d,960)
13. (220.1,1182)
14. (220.1d,960,1182)
15. (533.6,398)
16. (196.2,122)
17. (531.3, Figs. 317, 318)
18. (411.5; 452.8,23 & Figs. 2,24,25; 495.65,141; 533.6,333,398)
19. (158.3,149,Fig. 94; 533.8,9)
20. (508.7,495)
21. (237.6)
22. (533.6,374; 533.8,220)
23. (399.3,699); See also (399.3,629,Fig. 298b).
24. (188.65,299)
25. (525.3,754,Fig. 605)
26. (534.6)
27. (475.6)
28. (87.6,406)
29. (414.9)
30. (34.4; 87.6,406)
31. (443.6,32)
32. (393.3,103). See also (67.4).
33. (235.4,44)
34. (452.4; 770.4)
35. (560.4)
36. (560.4)
37. (540.3,97-98)
38. (540.3,96)
39. (525.3,579)
40. (235.4,44; 235.7,113,115)
41. (334.58,143); Dixson (140.1) says the spider monkey (Ateles) also has a baculum.
42. (525.3,602)
43. (259.8,323-324)
44. (158.3,209)
45. (405.9,376-377)
46. (235.4,44)
47. (292.6)
48. (495.063)
49. (235.7,115). See also (348.2; 452.8,88a).
50. (196.2,1300; 525.3,602)
51. (235.4)
52. (196.2,1300)
53. (385.5,63)
54. (490.3,336)
55. (583.3)
56. (399.1)
57. (140.1,149)
58. (334.6)
59. (245.1,200; 485.2)
60. (485.2)
61. (100.1,12; 231.88)
62. (225.4,162)
63. (385.5,79)
64. (385.5,63)
65. (101.1,13)
66. (205.1)
67. (205.1,194)
68. (405.9,376,Fig. 543, 485.2)
69. (235.7,116)
70. (53.9,250; 235.7,116)
71. (325.8,209)
72. (460.5,72,Table 3)
73. (245.1,207; 405.5,II,380; 588.4,1177)
74. (360.7,108; 490.3,Plates 1-5; 495.65,142)
75. (220.1d,1025-1026)
76. (235.1,134-135). See also (533.6,420).
77. (405.6,377)
78. (530.9,Fig. 250)
79. (505.1). See also: (495.06,266; 495.65,139-140).
80. (235.4,32)
81. (405.9,377,Fig. 545)
82. (503.3,471)
83. (74.7,60)
84. (503.3,471)
85. (198.3)
86. (187.6,412)
87. (139.6)
88. (192.2,115; 230.6,39; 231.82; 273.2; 603.1,30; 603.2,299b)
89. (139.6,192)
90. (36.4,386-387)
91. (200.1,210)
92. (460.5,87)
93. (100.1)
94. (316.7)
95. (100.1,11)
96. (100.1,11)
97. (100.1,11)
98. (460.5,72)
99. (588.4)
100. (23.4; 205.1; 205.4; 558.8)
101. (205.1,198)
102. (205.1,App. D)
103. (205.1,132)
104. (588.4,1175)
105. (443.6,276)
106. (540.3,97-98)
107. (74.3)
108. (167.5)
109. (73.3)
110. (246.5)
111. (102.1)
112. (589.4)
113. (544.3)
114. (235.4,44)
115. (534.1)
116. (573.3)
117. (191.6)
118. (168.8,200-201)
119. (194.3)
120. (168.8,201). These authors refer to Straus’s comments in (540.3).
121. (136.9)
122. (533.6; 533.8)
123. (259.8)
124. (540.3,97)
125. Ibid.
126. (533.6,399)
127. (544.3)
128. (334.4)
129. (242.7)
Most shared on Macroevolution.net:
Human Origins: Are we hybrids?
On the Origins of New Forms of Life
Mammalian Hybrids
Cat-rabbit Hybrids: Fact or fiction?
Famous Biologists
Dog-cow Hybrids
Georges Cuvier: A Biography
Prothero: A Rebuttal
Branches of Biology
Dog-fox Hybrids