On the Origins of New Forms of Life
9.2: Synapsids
(Continued from the previous page)
|
|
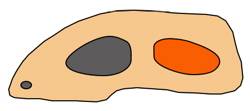
Synapsids had skulls with one temporal opening.
Credit: Buhler |
|
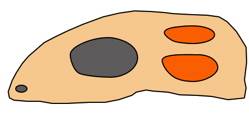
| A dyapsid skull has two temporal openings. Credit: Buhler |
{kind=link}
{kind=link}
Synapsids were creatures that are often described as "mammal-like reptiles." They could just as reasonably be called "reptile-like mammals" because they exhibit both reptilian and mammalian traits. Whatever they were, these animals had their heyday in the Permian Period (286–251 mya), right before the Mesozoic Era.1 They derive their name from the fact that they had a single temporal opening on each side of the skull, as is the case in mammals.
Among paleontologists, the tendency is to call animals with a single such opening "synapsids" if they are older than the Jurassic (before about 206 mya) and to call them "mammals" if they are younger. For this reason, the "origin of mammals" is more a verbal convention than a precisely dated historical event. As Nowak (1999: 1) points out,
Many synapsids had features considered characteristic of mammals, not reptiles, such as the presence of two occipital condyles (bony knobs forming a joint between the head and neck) and well-differentiated teeth.2 But the these animals have traditionally been classified as reptiles, apparently, for theoretical reasons since standard accounts of the history of life say
- that mammals evolved from the synapsids;
and - that mammals evolved from reptiles.3
If both of these assertions are assumed to be true, then synapsids have to be classified as reptiles.
What were synapsids?
But were synapsids conventional reptiles? Or were they a distinct type of animal that arose concurrently with reptiles? The earliest known synapsid fossils are as old as the earliest known reptile fossils. As Romer notes, "Primitive synapsids were already present in the Pennsylvanian — perhaps even the early Pennsylvanian." The Pennsylvanian began more than 300 million years ago, which means the synapsids long antedate dinosaurs, pterosaurs, ichthyosaurs, and all the other "saurs" that have come to characterize the Mesozoic Era.

|
|
Skull of Kannemeyeria erithrea
(Enlarge image) |
|
|
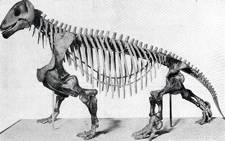
The Permian herbivore Moschops |
|
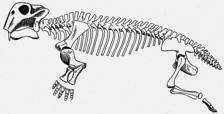
| Lystrosaurus |
Certainly, many different kinds of synapsids are known. There were herbivorous, insectivorous, and carnivorous forms, some fairly large. The early Triassic (approx. 230 mya) herbivore Kannemeyeria was the size of an ox.4 Another, earlier herbivore, Moschops of the late Permian (approx. 260 mya), was even bigger — 16 feet long.5 Contemporary with Moschops were smaller, doglike, carnivorous synapsids (Lycaenops) with long canine teeth and running legs. These animals are believed to have hunted the large herbivores in packs, like wolves.6 Lystrosaurus was a hippo-like aquatic creature with a widespread distribution7. The mole-like insectivore Cistecephalus lived underground, where it probably dug for bugs and worms.8Estemmenosuchus was a late Permian carnivore whose skull had strange bony protuberances, which may have borne horns in life.9 The tritylodonts of the late Triassic and early Jurassic10 were small herbivores with rodent-like teeth.11
An important evolutionary question, then, arises: Did all these varied types simply die out, as modern dogma would have it? Or are the various extant kinds of mammals descended from similar synapsid types?
The Problem of Diversity. Neo-Darwinian theory holds that all the functional diversity, evident in the various synapsid types, was in fact wiped out as Mesozoic reptiles usurped the synapsids' "adaptive niches." Typically, the claim is made that only the "basic structural plan" of mammals was derived from the synapsids. This view is somewhat peculiar because, in terms of bony remains — the only basis on which the basic structure of an extinct animal can be reconstructed — few traits consistently distinguish mammals from reptiles. In fragmentary fossils, even these traits are largely undetectable. But even when they can be discerned, we have already seen there is no reason to immediately conclude that such traits were associated in life with the various non-fossilizable traits defining mammals and reptiles. What, then, exactly is the "basic structural plan" of a mammal?
The notion of a basic structural plan seems to reflect only an attempt on the part of paleontologists to accommodate a constraint imposed by theory, which demands a common ancestor. In the minds of many biologists, the common ancestor is a pliable entity representing a general taxonomic category in its "primitive," incipient state (this is the "archetype" discussed in Section 8). Neo-Darwinian theory says that with the passage of time natural selection shapes this general entity (which has, at the outset, the full set of traits that define its corresponding category) into a variety of specialized forms suited to various niches (but still belonging to that category). This pattern of shared descent from a single common stem form, the key evolutionary topology of neo-Darwinian theory, is termed monophylety. In contrast, when new types arise from similar old types (i.e., new carnivores from earlier carnivores, new herbivores, from earlier ones), the pattern is "polyphyletic" — there is no common stem form. If stabilization processes involving hybridization are common modes of evolution, then polyphylety is a widespread pattern.
Orthodox theory usually says the first mammal came into existence about 220 million years ago. Most commonly, the morganucodonts are accorded this honor. They are typically described as "small, primitive, generalized" animals. It is thought they were insectivores. All later mammalian orders, both extant and extinct, are supposedly derived from this common ancestor (monophylety). Only one line of descent, passing through the pantotheres of the late Jurassic (approx. 150 mya), is supposed to lead from these "first mammals" to modern marsupial and placental mammals. Theory says all other Mesozoic animals categorized as mammals, and not in this line, died out without issue — and that any special traits they might have had (horns, gnawing teeth, hooves, and so forth) disappeared with them.12 The diversity of form present in these other types, and their presumptive suitability for a variety of ways of life, are generally ignored because it is assumed their lines ended in oblivion. But such conclusions are in no way necessary, nor, after careful consideration, are they even plausible. An evaluation of the probable origins of two extant mammals, the armadillo and the pangolin, will begin to reveal the fallacy of this mode of thought.
1. Early synapsids are usually called pelycosaurs; later ones, therapsids.2. Romer (1966).
3. For example, Stokes (1982: 362).
4. Case (1934).
5. Dixon et al. (1988: 189); Gregory and Broom (1926).
6. Colbert (1948); Dixon et al. (1988: 189).
7. Strickberger (2000).
8. Dixon et al. (1988: 192); Kemp (1982: 138).
9. Kemp (1982: 86, Fig. 29).
10. For example, Beinotherium and Oligokyphus.
11. Dixon et al. (1988: 193); Romer (1966: 185).
12. Under the neo-Darwinian view, the only generally recognized exception to this claim is a separate line leading to the extant monotremes (the echidnas and duck-billed platypus).