
Mitochondrial DNA (mtDNA)
Analysis of available data suggests humans are derived from an ancient cross involving a female chimpanzee
|
A diligent scholar is like a bee who takes honey from many different flowers and stores it in his hive.
—John Amos Comenius
|
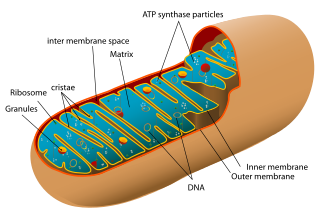 Cut-away view of a mitochondrion, showing its various contents, including mitochodrial DNA (mtDNA)
Cut-away view of a mitochondrion, showing its various contents, including mitochodrial DNA (mtDNA)
Have humans slowly diverged from chimpanzees, as standard evolutionary theory proposes? Or are we derived from ancient hybridization between chimpanzees and some other type of animal? An evaluation of mitochondrial DNA (mtDNA) suggests the latter, though, in itself, it tells us nothing about what that other animal might be.
For those readers who do not know what mitochondrial DNA might be, an explanation is perhaps in order. A living cell contains a variety of separate structures, termed organelles, each serving a specialized function (more about cell structure). Some of these organelles, for example the nucleus and the mitochondria, contain DNA. The nucleus has the lion’s share of the DNA because it contains the chromosomes, while the mitochondria have much less.a A mitochondrion (sing. of mitochondria) is shown in the figure above.
A well-known fact about mtDNA is that it’s maternally inherited, that is, offspring receive it from their mother only. The father makes no contribution. So in a hybrid cross involving a female chimpanzee, all of the hybrid offspring would have mtDNA identical to that of the chimpanzee mother. And if those hybrids, in turn, went on to mate among themselves to produce a race of descendants (in this case, the human race), the mtDNA of every descendant would also be identical to that of the original chimpanzee Eve, with the exception of the various rare mutations that would subsequently arise slowly over time.
If backcrossing of the hybrids to chimpanzees occurred in generations subsequent to the original cross but before the nascent human populations got established (the scenario proposed on this website), there still would be no contribution from the father participating in the original cross. All of the mtDNA in descendant humans would come either from our simian Eve or from any chimpanzee females participating in the ensuing backcrossing process.
With these basics out of the way, let’s look at some interesting facts.
Inferences from mitochondrial DNA
Certain studies investigating human and chimpanzee mtDNA have produced information that is difficult to account for under a simple divergence scenario. Thus, Davidson (1986, p. 697) points out that
In other words, if a randomly selected nuclear-encoded protein—say, alcohol dehydrogenase—is examined in a human and in a chimpanzee, it will be five times as likely to be present in two distinct forms in the human. Heterozygosity is a commonly used measure of genetic variability. If humans and chimpanzees were descended from a single ancestral population, mutation would have greatly increased the level of human nuclear variability relative to that of chimpanzees during the time since the two diverged. But in animals, mtDNA is believed to accumulate mutations more rapidly than does nuclear DNA (Brown et al. 1979). Therefore, if human nuclear variability has increased relative to that of the chimpanzee—which is the case—then human mitochondrial variability should have increased relative to the chimpanzee at least as much, if not more. The actual observation, however, is that chimpanzees have “three to ten times” the mitochondrial variability of humans (Wilson et al. 1985).
“The low level of mtDNA variability in humans fits with geological evidence that our species is a young one,” according to Ferris et al. (1981, p. 6322), who first noted this phenomenon. Wilson et al. (1985) have tried to explain this lack of human mitochondrial variability, saying it “could be indicative of a bottleneck” in the course of human evolution; that is, they suggest that the mtDNA present in modern humans is derived not from the prehistoric human population as a whole, but from a very small subset of that population. They think that all modern mtDNA can be traced back to a single female in the ancient population., or a few different females with the same mitochondrial type (ibid.). Only women come into consideration because, as has already been mentioned, mitochondrial genes are inherited from the mother alone.
This proposition has been referred to as the Eve Hypothesis. Gould (1987, p. 18) calculated that this “woman” may have lived in Africa sometime between 140,000 and 290,000 years ago.c Stoneking, et al. (1992, p. 174) used two different methods to date this common ancestor. One estimate yielded 133,000, the other, 137,000 years ago. These authors note that these dates come “close to the earliest dates ascertained for fossils of anatomically modern humans, raising the intriguing possibility that the human mtDNA ancestor might actually have been one of the first anatomically modern humans.”
It is difficult to reconcile such ideas with the fact that chimpanzee mtDNA, at least that of some chimpanzees, is virtually indistinguishable from that of human beings. As evidence of this fact, I quote in its entirety an abstract of a study carried out by Wesley Brown and Richard Hallberg (1972):
And yet, Hoyer et al. (1972) and Kohne (1970, Table 4) were both able to distinguish human nuclear DNA from that of chimpanzees by thermal absorbance melting alone. Thus, it appears that human mtDNA falls within the chimpanzee range of variation. If all humans are descended from a recent Eve, as Gould and Wilson suggest, then it seems that the chimpanzee providing mtDNA for Brown and Hallberg’s study was also descended from this female. But such a conclusion would be entirely inconsistent with conventional evolutionary thought. The usual assumption is that the human line separated from the simian millions of years ago, and that these two lines have evolved gradually in isolation up to the present. The idea that this divergence might have occurred as recently as one or two hundred thousand years ago is out of the question when theory is confined within conventional bounds.
It seems it was this obvious inconsistency that led Brown and Hallberg not to publish their findings in a full-length report. (The abstract above was of a talk presented at a conference. No formal paper followed.) When Brown came to visit as a seminar speaker at our department at the University of Georgia in the 1990s, I asked him why they had never published a paper based on these surprising results. Apparently, they were too surprising! He told me they had been convinced there was contamination (i.e., that they had somehow mixed up their samples and mistaken human mtDNA for chimpanzee mtDNA) because chimpanzee mtDNA could not possibly have been indistinguishable from human mtDNA. True. If you believe the standard story of evolution that humans “diverged” from chimpanzees millions of years ago, their results certainly would not have been possible.
Such difficulties do not arise under the assumption that humans are derived from hybridization between chimpanzees and some other kind of animal. If a female chimpanzee mated with a male that was not a chimpanzee, the hybrid offspring would have exactly the same mtDNA as the mother chimpanzee, but different, hybrid nuclear DNA derived in part from the “father’s side of the family.” A hybrid race descended from such a mating would have highly invariant mtDNA (which might very well be similar to that of certain living chimpanzees because the same mitochondrial type could have survived in both humans and chimpanzees, and thus be one of the extant chimpanzee varieties). But the intense mutation process (chromosomal recombination and restructuring), which would occur during hybrid meiosis in the generations after the cross, would increase nuclear variability beyond that seen in chimpanzees (meiosis has no mutational effect on mtDNA).
An extreme bottleneck involving human beings alone would, of course, have the same effect on mtDNA variability. But imagine the effect on human diversity if a single female were selected today to mother a new human race. The variability would go to nearly zero. Why, then, would our nuclear genes in actuality be five times more variable than those of chimpanzees when there is no evidence that chimps passed through a similar bottleneck? And why is our mtDNA indistinguishable from that of certain chimpanzees? These findings are consistent with the idea that humans are descended from a cross between one or more female chimpanzees and one or more non-chimpanzee males.
In short, Brown and Hallberg's findings would have been tantamount to proof that humans are hybrids descended from a cross involving a female chimpanzee, if they had not thrown their data away.
Notes:a. The lower end of the range for the former in animals is 6 x 107 nucleotide base pairs (bp), while the upper end of the range for mitochondria is only 106 bp (145.1,74). The typical mammal has more than 109 bp of nuclear DNA (305.6,73).
b. nuclear-encoded: the information required for making a “nuclear-encoded” protein is stored in genes contained in the cell nucleus (as opposed to mitochondrial genes).
c. This range is based on an estimated mtDNA mutation rate of between 2 and 4 percent per million years.
Most shared on Macroevolution.net:
Human Origins: Are we hybrids?
On the Origins of New Forms of Life
Mammalian Hybrids
Cat-rabbit Hybrids: Fact or fiction?
Famous Biologists
Dog-cow Hybrids
Prothero: A Rebuttal
Branches of Biology
Dog-fox Hybrids