
The Hybrid Hypothesis
3: An Initial Analysis
|
Then felt I like some watcher of the skies
When a new planet swims into his ken.
—John Keats
|
(Continued from the previous page)
|
I immensely enjoyed the piece on human origins. Clearly well-researched, painstakingly put together, and very convincingly written. I’ve shared a bunch of its more striking points with my girlfriend, who is Chinese (I'm presently in China), and whose diet contains a lot of pork, as is customary in China. She reports that she’s starting to feel pretty bad about eating pigs.
—Chase Dumont
|
While some domestic pigs are bred to be relatively hairless, all the wild pigs seem to be fully-furred. In fact, when domestic pigs go feral, they seem to immediately revert to a hairy form. If so, how could we have inherited the hairless trait from pigs?
When a pig escapes from a farm and starts living in the woods it does not suddenly become a hairy animal. Its descendants can, if they interbreed with hairy wild animals, but not otherwise. True, the Eurasian wild boar is hairy (though its hair is nowhere near as dense as, say, that of a cow or sheep). But we do not know the history of the domestic pig. It’s usually treated as conspecific with the Eurasian wild boar, but the two differ in chromosome counts (for the domestic 2n=38, but for the wild boar 2n=36). So it may be that they are not the same animal and that relatively hairless pigs similar to the domestic pig existed anciently. It may well be that the two have been treated as the same species merely because it has long been known that they can produce fertile offspring together. But these offspring may simply represent hybrids (this is one of many examples, by the way, of animals with differing chromosome counts producing fertile offspring together). The domestic pig has also hybridized with a variety of other types of pigs, but that does not imply that they are the same animal. For example, in addition to the wild boar, the domestic pig has hybridized with the Babirusa, Babyrousa babyrussa (pictures); Bush Pig, Potamochoerus larvatus (pictures); Bearded Pig, Sus barbatus (pictures); Visayan Warty Pig, Sus cebifrons (pictures); Sulawesi Wild Boar, Sus celebensis (pictures); and probably Sus oliveri and Sus philippensis. So why assume that the domestic pig and wild boar are the "same" animal? Relatively naked animals similar to the domestic pig might have existed anciently. We don’t really know what pigs looked like thousands of years ago, but a prehistoric painting in Altamira Cave in Spain shows a pig (pictures) that looks fairly naked, except in a few hairy regions (beard, scalp, neck and shoulders).
Video: A bearded pig
Some of the most easily accessible evidence that can be used to evaluate the hybrid hypothesis is visible in the mirror. In this section, we will consider certain external features that link humans with pigs. Much of my research on pigs has centered on the ordinary pig (Sus scrofa). Of course, ordinary pig is really a catchall term for a variety of breeds. "There are currently some 87 breeds of domestic pigs in the world, most of them in Europe and North America," according to Pond and Houpt, and "another 225 or more groups of pigs not recognized as breeds but each having unique characteristics, appearance, or geographical location."1 However, the focus here will be on traits that are generally characteristic of Sus scrofa.
And now, let’s look a little more closely at some human distinctions that, as it turns out, are characteristics of pigs as well. Traits that distinguish us from chimpanzees and other primates are the only ones that will be discussed, because traits that humans share with primates have no bearing on the question of whether humans are of hybrid origin. Under the hypothesis being considered, it would make no difference if humans are more similar to chimpanzees in most respects than to pigs. The interesting finding is that those features that do distinguish humans from chimpanzees and other primates can be consistently accounted for by reference to the pig.
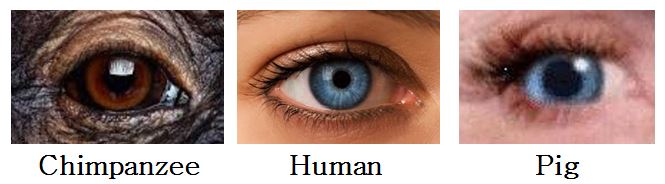
This physical affinity of humans and pigs is easily observable in certain external features. This fact did not escape Thomas Mann (1955, p. 271), who once wrote "The pig with its little blue eyes, its eyelashes and its skin has more human qualities than any chimpanzee — think how often naked human beings remind us of swine." Although I do not concur in Mann’s assertion that pigs share more traits with humans than do chimpanzees, I do think pigs and humans share more than enough traits to suggest a relationship. For example, lightly pigmented eyes, in shades of blue, green, and tan, are never found in chimpanzees or orangutans (Schultz 1947, p. 11). There is, apparently, only one known case of a gorilla with blue eyes (Dixson 1981, p. 31). Light-colored eyes are also rare in other primates (ibid.). Why, then, are they common in certain human populations? Where did this trait come from?
One possible explanation is that it was inherited from blue-eyed pigs. Blue is a common eye coloration in swine (as are green, yellow, and tan). The dark pigment (melanin), found so consistently in the irises of nonhuman primates, is beneficial. It absorbs ultraviolet light. To protect their eyes from these damaging rays, pigs depend on their narrowly slit, heavily lashed eyelids. Humans shield their eyes in a similar way, unlike the typical wide-eyed, sparsely lashed ape. [A reader, by the name of Chase Dumont, wrote in with the following comment, which is of interest in the present context: "The outer appearance of the eye itself looks quite different from a chimpanzee’s and more like a pig’s — the pupil/iris in a chimpanzee eye covers the entire eye, while the pupil/iris in a pig eye occupy a much smaller footprint, displaying much of the 'white' of the eye — as in humans)."]
Neither is it clear how a protrusive cartilaginous nose might have aided early humans in their "savanna hunter lifestyle." As Morris (1967, p. 67) remarks, "It is interesting to note that the protuberant, fleshy nose of our species is another unique feature that the anatomists cannot explain." This feature is neither characteristic of apes, nor even of other catarrhines (Hill 1966, p. 29). Obviously, pigs have a nose even more protuberant than our own. In a pig’s snout, the nasal wings and septum are cartilaginous as ours are (Walker 1983, p. 1175). In contrast, a chimpanzee’s nose "is small, flat, and has no lateral cartilages" (Sonntag 1923, p. 397). A cartilaginous nose is apparently a rare trait in mammals. Primatologist Jeffrey Schwartz goes so far as to say that "it is the enlarged nasal wing cartilage that makes the human nose what it is, and which distinguishes humans from all other animals" (Schwartz 1987, p. 185). The cartilaginous structure of the pig’s snout is generally considered to be an "adaptation" for digging with the nose (rooting). Rooting is, apparently, a behavior pattern peculiar to pigs. Other animals dig with their feet.
A protruding nose is perhaps the most prominent difference between a human face and that of a chimpanzee, but discussions of human evolution rarely mention the nose, perhaps because its lack of utility precludes explanation in terms of adaptation. Instead, most analyses deal with the fleshless skull, where the protrusiveness of the human nose is a bit less obvious (but visible nonetheless). It is a peculiar omission, because useless (nonadaptive) traits are widely considered to be the best indicators of relationship. What is the evolutionary utility of our unique nasal structure? Is it functional? Or is it the genetic residue of an ancient hybrid cross?

Another feature to consider is the philtrum, the dent seen on the center of the human upper lip. Apes lack this typical human feature.11 It seems a useless structure from a survival standpoint. Why is it seen, then, the world over in Homo? In both human beings and pigs, during the early stages of development, the upper lip is cleft, though I have not been able to find any evidence of such a cleft in the embryos of any nonhuman primate. As development continues, this cleft usually closes in humans, but persists in pigs.12 The human philtrum is a visible residue of this primordial split lip. In those human beings where this split never closes, the condition is known as cleft lip, a common birth defect. The frequent occurrence of cleft lip in humans is hard to explain if it is assumed that we are closely related only to primates. If the assumption, however, is that human beings are derived from a pig-chimpanzee cross, this finding becomes far more understandable.
@Macroevo Honestly, the pig-chimp hypothesis is simply genius. Applied common sense that makes me wonder how nobody realized it. Amazing
— alfredo romeo (@aromeo) September 1, 2013
Similar thinking explains the shortness of the human upper lip (distance between mouth opening and nostrils). Why has our upper lip become shorter and thicker in the course of evolution? All apes have upper lips much longer than those of humans,13 but a pig’s upper lip is so short that it is scarcely more than an appendage of the snout.14 Morris15 makes much of the fact that human lips are covered on their exterior surface by glabrous (i.e., absolutely hairless) mucous membrane:
He goes on to suggest that our peculiar lips are the product of "sexual selection." But other explanations are conceivable: In describing the skin of pigs, Getty16 states that "there are no true glabrous surfaces other than the labial borders," which are composed of red mucous membrane.
In reference to human earlobes, Morris observes that "anatomists have often referred to them as meaningless appendages, or `useless fatty excrescences.' By some they are explained away as `remnants' of the time when we had big ears. But if we look to other primate species we find that they do not possess fleshy earlobes. It seems that, far from being a remnant, they are something new."17 Perhaps, however, they are really something old on a new face. Sisson describes the lower portion of a pig’s ear as "strongly convex below, forming a prominence somewhat analogous to the lobule of the human ear."18
An additional feature of the human ear should be mentioned here, the Darwinian tubercle (see Darwin’s illustration below). In his Descent of Man, Darwin comments on this feature sometimes found on the rim of human ears which he describes as "a little blunt point, projecting from the inwardly-folded margin, or helix … These points not only project inward, but often a little outward, so that they are visible when the head is viewed from directly in front or behind. They are variable in size and somewhat in position,

Darwinian tubercle
(Darwin, 1871)
Primatologist Adolph Schultz (1973), however, flatly contradicts Darwin, saying that "clearly pointed ears, commonly called `satyr ears,' are among monkeys typical for only macaques and baboons and do not occur in any hominoids [great apes], not even in the early stages of development. There is no justification, therefore, to interpret the occasional `Darwinian tubercles' on human ears as an atavistic manifestation of ancestral pointed ears."20 But Schultz has not, perhaps, taken into consideration the pointed ears of swine.
Swine have prominent eyebrow hair. On the brows of the chimpanzee fetus it is possible to discern a region of light-colored bumps following a pattern similar to that of the human eyebrow. Adult apes, however, have no eyebrow hair.21 On their eyelids, pigs have luxuriant eyelashes, thicker even than those of human beings. In many pigs these cilia, as anatomists term them, are so thick that the animal seems to be wearing false eyelashes. But apes scarcely have eyelashes at all, despite the apparent survival value of this feature. Also, pongids have prominent brow ridges while pigs and most humans do not. If we choose to explain the development of human eyelashes and eyebrows in terms of natural selection, we must wonder why apes, which have existed at least as long as any hominid, have failed to acquire them. Perhaps their heavy brow ridges sufficiently protected their eyes, but if such is the case, why did not brow ridges also suffice for Homo? What was the pressing need that caused Homo to substitute tufts of hair for ridges of bone?
Dermal Characteristics
Update (8/14/2024): Most of this webpage was written more than ten years ago. Recently , I reported the results of a large-scale genetic study providing hard DNA evidence consistent with the hypothesis that humans are derived from a pig-bonobo cross (and moreover, inconsistent with the idea that we have gradually diverged from the apes). You can read a full report here. Alternatively, you can listen to an audio summary here. So it would seem that the difficulties described below (this section was previously titled "Why it may not be easy to evaluate this hypothesis with genetic data") have now been overcome.
In connection with the hypothesis that human origins can be traced to a hybrid cross, it’s important to realize that in most mammalian hybrid crosses, the male hybrids are usually more sterile than are the females. This fact means that breeders working with hybrids typically mate fertile females with one of the two parents (that is they "backcross" them). They do not, as a rule, produce new breeds by breeding the first-cross hybrids among themselves.
Often, even after a backcross, the resulting hybrids are still fertile in only one sex. So repeated backcrossing typically occurs. However, after a sufficient number of backcrosses, fertile hybrids of both sexes are often obtained and the new breed can thenceforth be maintained via matings among the hybrids themselves. Repeated backcrossing tends to be more necessary in cases where the parents participating in the original cross are more distantly related and genetically incompatible. So one expects also, in the case of new types of organisms arising via natural hybridization, for backcrossing to be the usual route to fertility and reproductive stability. And the same would hold in the specific case of humans arising via hybridization.
However, backcross hybrids are hard to identify with molecular techniques, and they become more so with each successive backcross (Vähä and Primmer 2006, Engler et al. 2015). Vähä and Primmer say that to reliably identify even first backcrosses by conventional techniques, at least 48 loci must be examined (and with repeated backcrossing that number greatly increases). To better understand this problem, consider the hypothetical effect of repeated backcrossing on the human genome. The reader may not be familiar with the phenomenon of gene conversion, but its effect on hybrids during backcrossing is to quickly homogenize gene sequences. To understand why this is the case, consider the effects of backcrossing on hybrid DNA.
 A Holliday junction. During meiotic recombination, double strands of parental DNA, shown here moving into the junction, separate into two single strands. Each single strand from one parent then joins with a single strand from the other parent. The resulting composite double strands then move out of the junction, undergoing gene conversion in the process.
A Holliday junction. During meiotic recombination, double strands of parental DNA, shown here moving into the junction, separate into two single strands. Each single strand from one parent then joins with a single strand from the other parent. The resulting composite double strands then move out of the junction, undergoing gene conversion in the process.
In the figure above, note that in either of the parental (incoming) double strands each nucleotide in one strand is properly paired with its complementary nucleotide in the other strand, A is always paired with T, and C is always paired with G. So the paired double DNA strand from one parent might look like this:
AGTTCCGACGCG
TCAAGGCTGCGC
while the strand from the other parent might look like this:
AGCTCCGACGCG
TCGAGGCTGCGC
In each of these two double strands all nucleotides are paired with the complementary nucleotide (A always with T, and C always with G). But when one double strand is compared with the other it’s clear that they differ at the third nucleotide base position. In the first double strand the nucleotide base pair is T-A, while in the second it is C-G.
Thus, when these strands associate with their new partner strands after passing through the Holliday junction in meiosis, the two resulting double strands will be:
AGTTCCGACGCG
TCGAGGCTGCGC
and
AGCTCCGACGCG
TCAAGGCTGCGC
So T is paired with G in the first outgoing strand and C is paired with A in the second outgoing strand. Gene conversion converts each such mismatched pair into a matched one by replacing one of the two nucleotides with the complement of the other nucleotide (which remains unchanged). Experimental results suggest that the mechanism chooses at random which of the two nucleotides to replace so that the nucleotides derived from either parent survive at equal rates.
So, in any given case of backcrossing, suppose the genomes of the original parents A and B, which produced the first-cross hybrids, differed at one nucleotide position in five (20%). Then the DNA in gametes of the hybrids would differ from A at one position in ten and from B at one position in ten. That is, the first cross hybrid’s gametes would be right in the middle between A and B with respect to gene sequence.
However, after one backcross to A, and the resulting gene conversion during meiosis in the backcross hybrids, the gametes produced by backcross individuals would differ from A at only one position in twenty (5%). And gametes produced by second backcross hybrids would differ at only one position in forty (97.5% similarity). It’s clear then that it rapidly becomes quite difficult to distinguish, on the basis of nucleotide sequence data, the backcross hybrids from the pure parent A to which backcrossing has occurred. Chimpanzees and humans are about 98% similar in terms of their nucleotide sequences.
The specific genetic underpinnings of the many traits that distinguish humans from chimpanzees are explained, in terms of the present theory, in a separate section entitled Why are Humans Different from Chimpanzees?
That humans lack the hair cover of nonhuman primates is an accepted fact. "It is this single factor that constitutes the chief difference between human skin and the skin of other mammals" (Montagna22). Some writers say that the hair coat of a chimpanzee is "sparse." But if "sparse" describes chimpanzee pelage, then "naked" accurately describes the skin of human beings. Any human who even approached the hairiness of other primates would be considered abnormal. Pigs, however, are a different case. Many domestic pig breeds have skin just as naked as human skin. As Cena et al. (101.9,521) observe, "Hair densities [of animal coats] range from the sparse residual covering on man and the pig with 10-100 hairs per cm², to [the] dense coats of species such as the fox and rabbit with about 4,000 per cm²." In wild Sus scrofa, according to Haltenorth, the density of hair coverage varies from "sparse to thick," depending on the specimen or variety in question.23 For example, the hair of the modern day wild variety of Sus scrofa present in Sudan (S. s. senaarensis) is quite sparse.24
Other primates do not have the long mane of hair that tops the head of an unshorn human, nor do they have beards. Haltenorth notes that in some varieties of Sus scrofa, manes are found on the neck and back , beards on the cheeks, and shocks of hair on the forehead and atop the head. He also says that the last of these three traits is found, among pigs, in Sus alone.25 A prehistoric painting of a pig found in Altamira Cave in northern Spain depicts an animal with a beard and thick hair atop its head (pictures). Sus barbatus, an extant pig native to southeast Asia (which forms fertile hybrids of both sexes in crosses with S. scrofa) has little hair on its body, but does have a very thick and bushy beard.26
Panniculus adiposus. In an article on the evolution of human skin, renowned cutaneous comparative anatomist William Montagna (1985, p. 14) notes that, "Together with the loss of a furry cover, human skin acquired a hypodermal fatty layer (panniculus adiposus) which is considerably thicker than that found in other primates, or mammals for that matter. This is not to say that only man has a fat skin, but a thick fatty layer is as characteristic an attribute of human skin as it is of pig skin." Similarly, Dyce et al. (1987, p. 742) note that there is a "well developed fat deposit present almost everywhere in the subcutis." Primatologist F. W. Jones (1929, p. 309) also noted this fat layer:
Panniculus carnosus. "Another particularity of human skin is its general lack, or loss, of the cutaneous skeletal muscle layer (panniculus carnosus) found throughout the skin of most other mammals. Remnants of a panniculus carnosus in human skin are found at the front of the neck in the apron-like, thin platysma muscle … All other primates, even the great apes, have a panniculus carnosus over much of the body" (Montagna (1985, pp. 14-15; see also Sobotta 1983, figs. 459, 460). As in humans, the cutaneous musculature of pigs is well developed in the neck (platysma muscle) and face, but sparse or nonexistent elsewhere (Nickel et al. 1986, p. 230; Sack 1982).
In animals having a panniculus carnosus, the skin receives its blood supply from direct cutaneous arteries (large superficial vessels running parallel to the skin surface in the cutaneous muscle sheath). But when no panniculus carnosus is present, arteries feeding the skin rise up like little trees from deep within the body. Arteries of this latter type are called musculocutaneous. These two forms of dermal circulation are depicted in the illustration below. Both pig skin and human skin are supplied by musculocutaneous arteries (Montagna and Parakkal 1974, pp. 143-144). As Daniel and Williams (1973, pp. 20-21) observed in an article on skin flap transfer, "Most experimental animals do not have a vascular supply to the skin similar to that of man. The pig’s cutaneous vascular supply has been demonstrated anatomically and surgically to be more comparable than most to that of man … As in man, the pig’s skin is supplied by ubiquitous musculocutaneous arteries and by a few direct cutaneous arteries." This observation has been confirmed by other authors: "Except for pigs, whose cutaneous vasculature resembles that of man, loose-skinned mammals are vascularized by direct cutaneous arteries" (Montagna and Parakkal 1974, p. 144). Therefore, in this respect, human skin is more similar to pig skin than to that of nonhuman primates: "Actually, the vascularity of the skin of most nonhuman primates is essentially similar to that of other furred animals" (Montagna 1985, p. 16). In particular, Baccaredda-Boy (1964, p. 211), as well as Moretti and Farris (1963, pp. 162, 170-171) found that the skin of chimpanzees differs from that of human beings in having numerous large, superficial vessels (i.e., direct cutaneous arteries).
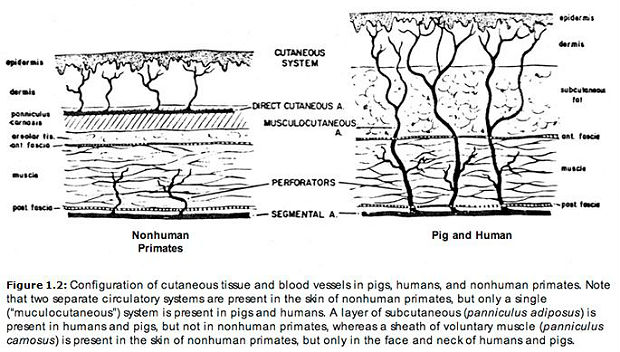
Human skin also stands apart from that of other primates — and from that of most other mammals for that matter — with respect to the quantity of blood that can be circulated through it (Montagna 1985). A certain amount of blood is needed just to feed the skin. This is the amount it receives in most animals. In humans, however, the maximum blood flow can be more than a hundred times greater than this minimum (Burton 1961; Montagna and Parakkal 1974, p. 151; Rowell 1977, p. 155). Fed by temperature-sensitive musculocutaneous arteries, the densely spaced cutaneous capillaries of human beings play an essential thermoregulatory role (Montagna 1985, p. 16). When the body begins to overheat, large quantities of warm blood can be rapidly cooled in these capillaries via sweat evaporation. One measure of cutaneous vascular density is the capillary loop separation interval. In human beings, the typical distance between capillaries ranges from 50 to 100 microns (Ryan 1973, p. 619). In porcine flank skin, this figure is reduced to only about 20 microns, a separation interval so small as to be almost incredible. When white pigs are exposed to high temperatures, the skin flushes pink with blood (even in the absence of sunlight) as it does in light-skinned human beings under similar conditions (Roller and Goldman 1968, p. 13).

|
Human flea, Pulex irritans |
Fleas. Perhaps this difference between our cutaneous vasculature and that of our primate kin, the richness of the feast beneath our skin, accounts for another human distinction: "Ironically," writes Nicole Duplaix (1988, p. 694), "man is unique among the primates in having fleas." More than 2,400 distinct types of fleas have been treated as species or subspecies (ibid., p. 689). Parasites are usually rather specific in their choice of host. Fewer than twenty of these 2,400 types will readily bite human beings (ibid., p. 685). Foremost among those that feed on Homo sapiens is the human flea, Pulex irritans, but we are not the only suitable hosts for this species. According to Bennett (1975, p. 775), "Pulex irritans, the human flea, breeds freely in hog-house litter and may become a serious pest of swine."
The panniculus adiposus replaces hair as an insulating layer in human beings and pigs. According to Beckett (63.8,2),
In the figure above, notice that the musculocutaneous arteries pass through the cutaneous fat. This perforated fat layer constitutes an insulating mechanism that can respond quickly to ambient temperature, a characteristic that hair lacks. Dilation of the musculocutaneous arteries in response to heat increases blood flow to the skin. This increase in circulation can raise skin surface temperature to a level almost as high as that within the body, thus increasing the rate at which heat is lost to the environment.b In cool environments, constriction of these arteries reduces skin temperature and, consequently, the rate at which body heat is lost to the atmosphere because the fat layer can then serve as an insulating blanket.
Possession of a panniculus adiposus allows adjustment to changes in ambient temperature on a moment-to-moment basis — a clear advantage in the temperate zones where much of the human race has made its home, because these regions are much more subject to sudden, extreme shifts in temperature than those close to the equator. Nonhuman primates and other furred animals do not have the option of adjusting their skin temperature. Because their skin is not insulated from the rest of the body by a layer of fat, its temperature must remain near that of the flesh beneath it.
Pig skin is separated from the inner body by a thick fat layer, and it can cool to an extreme degree. Fat, not hair, is the primary insulating barrier.47 Alaskan swine can withstand sub-zero temperatures by cooling their skin to as little as 9℃ (at an ambient temperature of -10℃) without suffering tissue damage.48 Acclimatized human beings, too, can reduce skin temperature to about 10℃ without injury.49 This mode of insulation is completely different from that of nonhuman primates, more like that seen in certain aquatic mammals (e.g., seal, walrus). With the exception of the pig, it seems that no other land animal has this form of insulation.
More than a naked ape, Homo is a variably insulated naked ape. In hot environments human beings (and pigs) can increase the circulation of warm blood to the skin and raise temperatures almost to the level of body core temperature, thus maximizing heat loss to the surrounding air. If weather turns cold, they can restrict cutaneous circulation, cooling the skin to such a degree that heat loss is significantly reduced. This ability is especially apparent in fat50 or acclimatized individuals.51 Although a cultural advance, the invention of clothing, made it possible for Homo to inhabit cool regions formerly off-limits to primates, a biological advance, in the form of a new insulation system, has increased the human ability to withstand the sudden temperature variations found in those regions.
Besides being a good insulator, human skin is surprisingly thick. "The epidermis over our general body surface ["hairy skin" see note at right] is substantially thicker than that of other primates: the horny layer [stratum corneum] can be peeled off intact as a diaphanous but tough membrane that can be used for experimental purposes … The epidermis in the hairy skin on nonhuman primates, mostly like that of any other furred mammal, is relatively thin, with a relatively thin horny layer" (Montagna52). Pigs, though, have a thick epidermis and stratum corneum, thicker even than that of human beings.53
The elasticity of our skin is also unusual. "Whereas the skin of the great apes and that of some of the simian primates have variable amounts of elastic fibers, in no animals, regardless of sex, age, or locality have we found the abundance of elastic tissue characteristic of human skin" (Montagna54). This finding comes from the same author who, in an earlier article comparing human skin with that of pigs, observed that "one of the most striking resemblances between these two skins [pig and human] is the large content of elastic tissue in the dermis."55
He also remarks that "the surface of both skins [human and porcine] is grooved by intersecting lines which form characteristic geometric patterns."56 In a separate paper on the evolution of human skin he provides a little more detail:
The presence of these lines in both pigs and humans is not easily explained in terms of natural selection since they have no known function.58
On the underside of our "hairy skin" (general body surface), where the epidermis meets the dermis, is a different patterning not corresponding in its configuration to the outside patterning described in the preceding paragraph. A similar, though coarser, pattern is also characteristic of the epidermal-dermal junction in pigs. Montagna, however, notes that "in split-skin preparations where the epidermis is neatly removed from the dermis, the epidermis of heavily haired animals is flat.59 Even in monkeys and apes, epidermal grooves are found only around the attachment of the ducts of glands and pilary canals." We can account for a finer patterning in humans than in pigs by the fact that a fine mesh is intermediate between the coarse patterning of pig skin and the smooth undersurface of nonhuman primate skin.
So, in the pig, we have a sparsely haired animal with a fatty, stretchy skin supplied by musculocutaneous arteries. The surface of the hairy skin is marked by congenital lines similar to those seen in human beings, and the patterning of the epidermal-dermal junction is also quite similar in the hairy skin regions. Under the hypothesis that we are considering, it makes little difference that pig skin differs from human skin in other ways. The essential point is that, in those cases in which our skin is peculiar for a primate, an explanation for each such anomaly can be found in the skin of pigs.
The Savanna Hunter
A mature pig has about 500,000 large sweat glands distributed over its entire body (503.3,497; 506.5,316). Nevertheless, it is often asserted in the literature that pigs do not sweat. This assumption can be traced to studies by Ingram and by Mount, who studied perspiration rates in immature animals, usually sedentary piglets (247.03; 247.1; 389.7; 390.1; 390.2; 390.3; 390.5). Studies evaluating pig sweating have concentrated on young pigs because they are of greater commercial interest. Immature animals are no more appropriate for determining the evaporative qualities of a boar or a sow than a toddler would be for revealing traits of an adult human—Children sweat much less than do adults (584.4,577). Small animals have a tendency to hypothermia (because their surface area is large in proportion to their size), not hyperthermia, and have little tendency to sweat (390.8,182). Perspiration in pigs is often overlooked because these animals are, apparently, more efficient sweaters than are humans. Their sweat glands seem to be better attuned to thermoregulatory needs (they produce no more sweat than what is necessary to cool cutaneous blood by evaporation). Very little sweat is lost to runoff because sweat rarely builds up on the skin. But observed rates of sweating in mature pigs are approximately comparable to those of humans. Beckett (63.4) found that a 350 lb. sow at rest lost approximately 95 g/m² in sweat per hour at a dry bulb temperature of 98℉ and wet-bulb temperature of 81℉. At a much higher temperature (122℉ dry bulb and 79℉ wet bulb), Myhre and Robinson found that 70 kg men at rest lost moisture (sweat + respiration) at a rate 250 g/m² per hour (398.7,Table 3). Even in smaller pigs (198 lb. gilts), skin moisture loss is important (387.8,Table 1), ranging from one-third to two-thirds of total moisture loss (lung + skin). The claim that pigs need a wallow when living in hot climates (because they supposedly do not sweat) is also encountered. But Heitman and Hughes exposed hogs without access to a wallow to high temperatures (100℉ relative humidity 35%) for a week without any fatalities—conditions where the only avenue for heat dissipation is evaporative cooling (232.5,176).
Pigs sweat when they are hot. "The apocrine [i.e., sweat] glands of the horse and pig secrete profusely during violent exercise and stress" (Montagna60). This sweating serves a thermoregulatory function in pigs just as it does in human beings.61 The hairy skin sweat glands of nonhuman primates, however, do not respond to thermal stimulation. The failure of nonhuman primates to sweat puzzled Montagna: "One might surmise," he writes,
In contrast, even a small dose of acetylcholine or adrenaline elicits sweating in pigs.63 Even the immature pigs studied by Ingram (247.1,95) responded to adrenalin.
The notion that nakedness has somehow enhanced sweat evaporation in humans is widely received. Supposedly, our sparse pelage allowed our ancestors to cool their skin more rapidly than hairy animals in hot, dry environments, or somehow improved their ability to dissipate metabolic heat while rushing about the savanna in pursuit of prey. Russell Newman, however, points out that our lack of reflective hair actually increases solar heat load and the need to sweat.164 To substantiate this claim, he cites a study by Berman showing that cattle exposed to the sun sweat more after their hair is removed.165 Similarly, panting increases in shorn sheep.166
Clothing, which replaces hair as a radiation barrier in human beings, has much the same effect on human perspiration. Human beings subjected to solar heat loads sweat more when naked than when wearing light clothing under otherwise identical circumstances. In a study of the effects of clothing on sweat, Adolph167concluded that "the nude man can save easily as much body water by putting on a shirt and trousers as can the clothed man by finding good shade." Moreover, body hair does not reduce convective heat loss "and has nothing to do with radiation of long-wave infra-red heat to cooler objects," says Newman.168 He therefore asserts that naked skin,
If increased radiant heat loads caused early humans to depend more on sweat for cooling, why has hair loss, which increases those loads, progressed to the degree that it has in Homo? Under the assumption that humans first evolved on the arid, sun-drenched savanna, it is difficult, in terms of survival efficiency, to account for a reduction in hair density that would result in increased rates of water consumption. Newman points out that there is no evidence that hair interferes with sweat evaporation. Actually, I myself performed a crude experiment, the results of which indicate that hair actually accelerates the evaporation of sweat. This finding is surprising in light of evolutionary theorists' frequent claims to the contrary. But with a little consideration, one realizes that a hair coat is not a vapor barrier. Fur’s ability to "breathe" has always distinguished it from less desirable insulators that slow heat loss but don’t "wick away" moisture from the skin. Why should hair not only allow, but even enhance, evaporation rates? There are at least two reasons. First, wet hair presents a more irregular surface to the surrounding atmosphere than does hairless skin, augmenting the surface area available for evaporation. Second, hair allows uniform dispersion of sweat by capillary action, preventing the formation of the individual droplets seen on naked skin. When such droplets form, the skin lying between them does not serve as an evaporative surface and the vaporization rate is reduced.
As the amount of sweat on the skin increases, the individual drops do merge to form a continuous sheet of water. But when a large amount of sweat is present on naked skin, another type of inefficiency sets in — runoff. More sweat runs off hairless skin without evaporating. The coat of a hairy animal acts as a sponge, retaining sweat in position until it can evaporate. Perspiration dripping off the body has no cooling effect, because no heat is absorbed by runoff. In contrast, evaporating sweat absorbs a large amount of heat. 1a But Adolph’s research indicates that about a quarter of human sweat can be lost to runoff, even under near optimal evaporative conditions. 1b A reflective hair coat, then, has three advantages: (1) lower solar heat loads; (2) increased rate of evaporation; (3) less sweat wasted on runoff. It is therefore difficult to understand how naked skin can be interpreted as an "adaptation" beneficial to a savanna hunter.
Of course, the "savanna hunter" hypothesis is just one of many theories. Hair loss in Homo has been the object of much speculation (for a survey of such theories, see 165.1). Besides those who say we lost our hair on the savanna170and/or because we were hunters,171there are others who suggest we may have lost it in the forest,172 or even in the sea.173 Some authors suggest that nakedness made us sexually enticing174or that hairlessness became thermally advantageous when we started wearing clothes.175
Even if we wished to assume that humans did at one time have a hair coat (there is absolutely no evidence that such was the case), these theories would not explain the advantage of a sparse coat of hair. The hunting hypothesis is untenable because nonhuman terrestrial predators all have thick hair coats. A similar objection can be raised to the sexual enticement scenario. Why haven’t all mammals lost their hair if nakedness is enticing? The aquatic proposal is also dubious, most small (human-sized or smaller) aquatic or semi-aquatic mammals do have hair coats.176
Hogu I
Amo an ape aghast
Hic haec hog hiccup
Pig latin lover
Hogu II
Amo amas amat
An ape aghast agape
Pig latin lover
Hogu III
An ape against a pig
Agape agape agape
Pig greek lover
Hogu IV
An ape agape and a pig
Again and again and again
Then a human
Hogu V
An ape agape
A pig and a poke
Vo-lar-e who-ho-ho-hog
Hogu VI
No pig no gain
Ape agape pig again
Pig in ape inhumane
Hogu VII
Pig in ape chasm
Fakes orgasm
Ham actor
Hogu VIII
Cheeky Chinese chimps
Like a pig in a china shop
Porcelain
The results of my evaporation experiment make it difficult for me to accept Mount’s opinion that naked skin evaporates sweat faster than hairy skin.177 For the same reason, I doubt Wheeler’s suggestion that the acquisition of erect posture by hominids "was probably the essential pre-adaptation which made it possible for them to shed body hair and develop extensive evaporative surfaces."178 Also dubious is Kushlan’s "vestiary hypothesis," because it proposes that the invention of clothing left Homo free to lose his body hair and thus obtain "the most efficient cooling system of any mammal."179 As we have seen, naked skin provides no particular evaporative advantage.
Because nakedness is a handicap on the savanna, Newman concludes, it is unlikely that human ancestors lost their hair after leaving the forest: "If one had to select times when progressive denudation was not a distinct environmental disadvantage, the choices would be between
Note, however, that Newman does not explain why our ancestors lost their hair in the same environment (forest) where apes did not. If humans came into being via hybridization between pigs and chimpanzees, their genesis would almost surely have occurred in the forest. Chimpanzees live in forests. On the basis of its high rate of water consumption, Yang concluded the pig, too, is functionally a forest animal.182 Human beings need more water than almost any other animal.183
Indeed, it seems incredible that a hominid would spend any more time than necessary away from the forest. Although the savannas of Africa were teeming with game, they were also swarming with ferocious predators. When a human being is chased by a lion, the first impulse is to find a tree. Consider the picture painted by current evolutionary theory: the noble savanna hunter, naked to the brazen sun, boldly erect on an arid and treeless plain, in indefatigable pursuit of a wary and dangerous prey, indifferent to the attack of rapacious carnivores. Certainly, this description has dramatic appeal. It’s like a Tarzan story. But is it plausible?
This ancestral shake-up is big.
Not just apes can lay claim to that gig.
I just figured out
Where I got my big snout.
Turns out great-granddad was a pig!
The above is a limerick read by Carl Kasell on National Public Radio’s Wait Wait... Don’t Tell Me!. Peter Sagal’s explanation: “Well, a geneticist trained at the University of Georgia has now theorized that the human race was born when a female chimpanzee got busy with a pig. And this is because humans and pigs share a startling number of traits, hairless skin, very rare in primates, light-colored eyes and a love of bacon.”
Excellent article: Georgia geneticist challenges evolution, links humans to pigs http://t.co/JIFFlR2JRs via @sharethis — luca turin (@lucaturin) July 29, 2013
|
CITATIONS:
1. (443.6,14)
2. (331.35,271)
3. (495.1,11)
4. (140.1,31)
5. (140.1,31)
6. (385.5,67)
7. (235.4,29)
8. (588.4,1175)
9. (533.6,397)
10. (505.1,185)
11. (533.8,103)
12. (530.9,Fig. 502)
13. (285.5,86)
14. (525.3,484)
15. (385.5)
16. (196.2,1421)
17. (385.5,66)
18. (525.3,923)
19. (135.6,I,21-22)
20. (495.9,52)
21. (158.3,124,161; 460.5,Plates 21,22,74; 533.6,415)
22. (360.8,109)
23. (230.25,24)
24. (153.4,173)
25. (230.25,4-5,Table 1)
26. (101.67)
27. (360.9,14)
28. (259.8,309)
29. (360.9,14-15). See also (531.3,Figs. 459,460).
30. (405.7,230; 485.2)
31. (360.85,143-144)
32. (125.1,20-21)
33. (360.85,144)
34. (360.9,16)
35. (50.1,211)
36. (379.9,162, 170-171)
37. (360.9,16)
38. (99.2; 360.85,151; 473.6,155)
39. (360.9,16)
40. (480.3,619)
41. (470.2,13)
42. (159.5,694)
43. (159.5,689)
44. (159.5,685)
45. (68.3,775)
46. (63.8; 390.2,293; 360.9,14; 447.2,77-78)
47. (390.2,293)
48. (250.2,418)
49. (250.8,183-184)
50. (329.5,70-71)
51. (250.8,183-188)
52. (360.9,7). See also (360.8,109; 360.85,29).
53. (90.1,485). See also (360.3,13,20,21).
54. (360.8,115).
55. (360.3,20).
56. (360.3,20)
57. (360.9,6)
58. (360.7,578)
59. (360.7,582)
60. (360.7,588). See also (63.4,165-166; 470.2,Table I).
61. (232.5; 360.7)
62. (360.8,122)
63. (178.5,199a)
164. (400.5,378)
165. (72.5)
166. (328.28)
167. (15.4,63)
168. (400.5,378).
169. (400.5,378)
170. See (118.5) and (595.4) among others.
171. (36.1; 277.4; 287.1).
172. (400.5,383)
173. (380.5)
174. (30.8,100; 385.5)
175. (286.3)
176. (595.4,24)
177. (390.8,42)
178. (595.4,26-27)
179. (286.3,29)
180. (193.4)
181. (400.5,382-383)
182. (602.9)
183. (400.5,379)