Hybrid crosses as probability distributions
|
We must not treat the unknown as known and too readily accept it; and he who wishes to avoid this error—as all should do—will devote both time and attention to the weighing of evidence.
—Cicero,
De Officiis, 1.6
|
The discussion in this section considers hybrid crosses as probability distributions. Under this heading there are four points of inquiry: (1) How can hybrid crosses be represented statistically? (2) How is the number of trials related to the probability of success? (3) What factors affect the likelihood of producing a hybrid? (4) How does survivorship differ in hybrid crosses? (5) How does the degree of difference of the parents participating in a cross affect the likelihood of being able to produce a hybrid?
1: How can hybrid crosses be represented statistically?
Hybrid crosses can be defined as matings between organisms that differ in some specified way. Each such mating can be viewed as an experiment in which a hybrid may or may not be produced. In the language of statistics, such an experiment is a binomial trial because it is a random experiment with exactly two possible outcomes, “success” and “failure,” in which the probability of success is the same each time the experiment is conducted. Call this probability p, and that of failure, q.
The value of p and q in any specific experiment will depend on circumstances. For example, in a mating between two types of mammals, the probability of producing a hybrid with a single insemination would depend on the volume of semen introduced into the reproductive tract of the female. The relation between p and semen volume might look something like this:
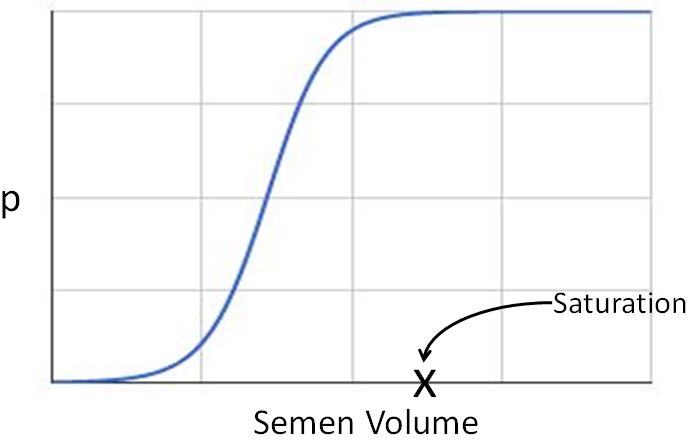
But for a given set of circumstances, the number of successes in n trials would be a binomial random variable with mean np.
How is the number of trials related to the probability of success?
If the binomial random variable X is the number of successful outcomes taking place during n trials, then the probability of at least one success, P(X≥1), approaches 1 (that is, certainty) as n approaches infinity. This is simple to prove. First consider that
P(X=1) = p+(1− p)p +(1− p)2p +⋯ = p/[1−(1− p)] = 1.
Now clearly, the probability that the event takes place at least one time is greater than it’s taking place exactly once, that is, P(X≥1) ≥ P(X=1). Therefore, given that 1 ≥ P(X≥1) for all values of n, P(X≥1) must also approach 1 as n approaches infinity.
A practical inference from the foregoing: Over evolutionary time, which allows for a near infinite number of trials, the probability of a hybrid cross successfully occurring at least once will be quite high even when p, the probability of success in any single mating, is very small.
3: What factors affect the likelihood of producing a hybrid?
Many different factors can affect the frequency with which a hybrid will be produced. Beyond the one already mentioned (semen volume), there are such factors as the number of separate inseminations carried out, the quality of the semen introduced, the age, health and fecundity of the female, the degree of difference between the two members of the mating pair, and so forth. For each of these factors, there will be some corresponding, specific probability of success, in a single trial where “success” might defined as the production of a hybrid, or as the production of a hybrid capable of producing offspring, or as the production of a hybrid of a certain sex, or color, or size, and so forth. The actual value of each such probability can be estimated only by observation and experimentation, as is the case with any other type of natural phenomenon.
4: How does survivorship differ in hybrid crosses?
In many hybrid crosses, there is a large increase in the number of early deaths, so much so, that such a finding might be said to be typical. Thus, frequency distributions for time of death from conception for a hybrid and a non-hybrid population would typically look like this:
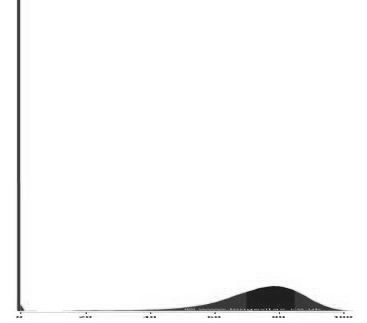 Typical pattern of death rates over time in a hybrid population (horizontal axis is time from conception, vertical axis is the death rate). Note the large spike in early deaths.
Typical pattern of death rates over time in a hybrid population (horizontal axis is time from conception, vertical axis is the death rate). Note the large spike in early deaths. Typical pattern of death rates over time in a non-hybrid population (horizontal axis is time from conception, vertical axis is the death rate).
Typical pattern of death rates over time in a non-hybrid population (horizontal axis is time from conception, vertical axis is the death rate).
5: How does the degree of difference of the parents participating in a cross affect the likelihood of being able to produce a hybrid?
Within the context of a hybrid cross, difference is often described in terms of taxonomic position, that is, a cross between parents classified as belonging to different genera, but the same family, is described as intergeneric. Similarly, a cross between parents classified as belonging to different families, but the same order, is described as interfamilial.
Now many biologists would say that hybrids can be produced from crosses that are no more distant than intergeneric, but that it is impossible to produce them from crosses that are interfamilial (or even more distant). This amounts to saying that:
- if the male and female participating in a cross belong to the same family, then a sufficient number of inseminations may produce a hybrid, but
- if they do not belong to the same family, then no matter how many inseminations take place, no hybrid will ever be produced.
So if these two assertions were true, a frequency distribution showing the effect of taxonomic difference on the probability of producing hybrid crosses might look like this:
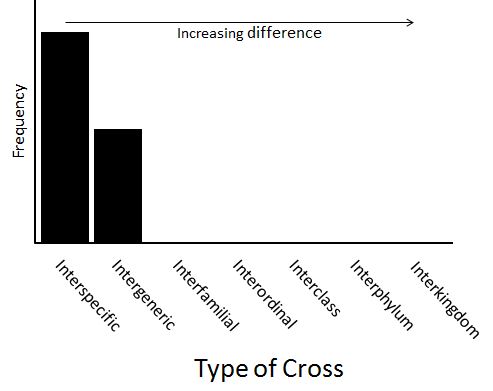
In other words, many biologists think that, for any cross that is more distant than intergeneric, the probability of occurrence is zero.
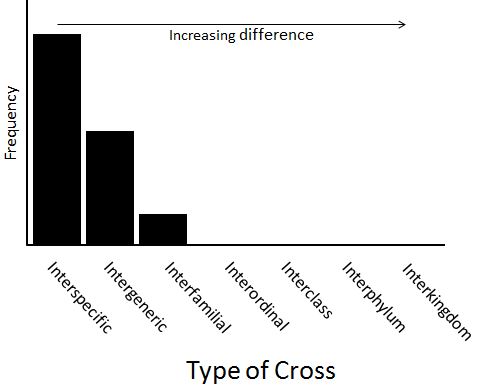
There are other, slightly more liberal-minded scientists, who say that interfamilial crosses occur, but certainly none that are more distant. Their view can be represented by the following frequency distribution:
 Pig-human hybrids
Pig-human hybrids Cow-human hybrids
Cow-human hybrids Sheep-human hybrids
Sheep-human hybrids Chicken-pigeon hybrids
Chicken-pigeon hybridsThat is, they say that the probability of occurrence is zero for interordinal crosses (and for any cross more distant than an interordinal cross).
However, because these two frequency distributions (that is, the two shown above) merely represent what scientists believe rather than what has been reported from observation, they are not scientific. As is the case with any other factor affecting the probability of producing a hybrid, the effect of degree of difference can only be determined in a scientific way via experiment and observation.
Though experimentation bearing on this topic has been limited, available observations suggest that interordinal and interclass crosses do occur, though at lower frequencies than less distant ones, that is, hybrids of this type seem to be rare rather than non-existent. For example, using artificial insemination, Newman (1915, 1917 and 1918) obtained interordinal hybrids from various fish crosses. Numerous interordinal and interclass hybrids are documented, too, on this website (see, for example, the articles linked from the images at right).
Another approach to developing a scientific answer to this question is to collect relevant literature such as breeders’ reports, zoo records, or accounts about unusual hybrids in books, magazines and newspapers. Many reports about interordinal or interclass hybrids are collected on this website (see the articles for interordinal and interclass crosses linked from this page).
Such preliminary results, then, would suggest that a frequency distribution showing the effect of difference on the probability of producing hybrid crosses might more accurately be represented by a frequency distribution like this:
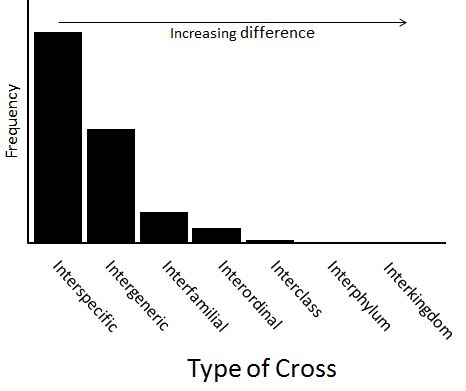
Obviously, more exact valuations of the class frequencies in this histogram will have to await more accurate observational results. But extrapolating from what is known, it seems likely that crosses even more distant (interphylum or interkingdom) probably occur as well, albeit at low rates.
A further point to mention in this connection is that, although the probability of success would be expected generally to decline with increasing taxonomic distance, the correlation is not strict. For example, when Terada et al. (2004; ||y3zgwhed) produced early-stage cow-human hybrids by microinjecting human spermatozoa into cattle eggs, 83% of the injected eggs became activated.
Moreover, the number of different types of distant crosses far exceeds the number of close crosses. For example, the number intrageneric crosses in which a given kind of organism can participate may be quite limited. Imagine a particular kind of organism a in a genus containing three kinds of animals a, b and c. In this case a can potentially participate in only two types of crosses, a × b and a × c. On the other hand, the number of interphylum pairings in which any given organism could potentially participate would be literally in the millions, which would tend to counteract the fact that the probability in any one of those many types of crosses would likely be minuscule.